FISICA DE PARTICULAS
OBSERVAR VIDEO Y ELABORAR RESUMEN.
http://www.youtube.com/watch?v=OtBqyehjcNs
FUNCIÓN CELULAR
- Resumen
- Estructura y funcion
celular:
- Organización y
procesos:
- Conclusiones:
- Referencias
bibliograficas
La célula es
la unidad constitutiva de la materia viva,
puede constituir por si sola un individuo o participar junto con
otros elementos semejantes en la formación de organismos
más complejos. La individualidad de la célula
está relacionada con su estructura y con la miscibilidad
con el ambiente de
las sustancias que la componen. Su estructura consta de algunos
elementos que aparecen diferenciados y constantes en cualquier
momento de la vida celular, excepto en el periodo reproductivo.
Para sus funciones y
desarrollo las
células
toman materiales
nutritivos del exterior, las sustancias nutritivas (y junto a las
mismas eventuales sustancias inertes o tóxicas) pueden
atravesar la membrana celular por fenómenos
fisicoquímicos o penetrar por sus poros.
Algunas células pueden realizar fotosíntesis, transformando la
energía luminosa en energía química, estas
células pueden ser algas verdeazules y
cianobacterias.
La
descripción de la división celular
para la comprensión de la misma, se la define como la
función
celular que garantiza la supervivencia de la especie mediante la
reproducción, por medio de la cual, una
célula es capaz de dar origen a dos o may células
hijas, semejantes a ella. Clásicamente, han venido
clasificándose a las formas de reproducción en las
siguientes variedades :la asexual (división directa,
gemación o abotonamiento, ivisión endògena,
carioquinesis (mitosis,
meiosis) y la
sexual (isogàmica y heterogàmiva).
En cuanto a la parte correspondiente a
microbiologìa no es posible hacer justicia a la
extraordinaria profusividad u variedad biológica de los
protistas; solamente unos cuantos representantes de cada uno de
los subgrupos han podido ser descritos de forma muy resumida. No
existe desgraciadamente un libro que
proporcione una visión màs extensa del grupo
biológico entero. La comprensión de la biología comparada de
los protistas eucariòticos se ve además impedida
por dificultades terminològicas, debido a que los
botánicos y zoólogos han aplicado nombres
totalmente diferentes a estructuras
comunes a los tres subgrupos.
DESCRIPTORES: Microbiologìa / Bacterias/
Hongos/
Protozoos/
Algas/Virus de animales y
vegetales.
2. ESTRUCTURA Y
FUNCION CELULAR:
-
La microscopía electrónica es una herramienta muy
importante en el estudio de la estructura celular. Su
principal ventaja reside en que proporciona imágenes de diferentes estructuras
celulares, en diferentes condiciones. Para determinar la
función de las estructuras celulares es necesario
introducir otras técnicas. Los investigadores deben
purificar distintas estructuras celulares de manera que se
puedan emplear métodos físicos y
químicos para determinar lo que cada una realiza. En
la actualidad los biólogos celulares emplean
técnicas experimentales distintas a comprender la
función de las estructuras celulares. (1)
-
-
La palabra célula proviene del
latín cellula = pequeña
estancia, unidad constitutiva del protoplasma o
materia viva; puede constituir por sí sola
un individuo o participar junto con otros elementos
semejantes, en la formación de organismos
más complejos. La teoría celular
sostenía que la célula era un
elemento estructural constante en todos los seres
vivos, desde los protozoos, constituidos por una
célula única, hasta los metazoos y
matafitos , animales y vegetales pluricelulares.
Sin embargo ciertas observaciones han disminuido el
valor de esta generalización;
se ha visto, por ejemplo, que no sólo no
pueden considerarse todos los protozoos como
verdaderamente unicelulares, sino que existen
organismos vivos carentes de cualquier estructura
celular, como los virus filtrables. También
las bacterias, aunque muestren algunas notas
estructurales cromáticas, no presentan un
verdadero y propio núcleo.
La célula típica, libre,
suele presentar forma esférica, y
esféricas son también las
células que flotan en los fluidos. Algunas
especies celulares tienen, por el contrario, una
forma propia, como los glóbulos rojos
ovalados de algunos anfíbios y mamíferos, y los
glóbulos rojos bicóncavos del
hombre.
La forma celular puede variar por la
acción recíproca de elementos,
formando colonias o tejidos, y depender también
de la diferenciación y de la función
de las mismas células
En cuanto a sus dimensiones, casi todas
las células son microscópicas: los
diámetros máximos varían desde
algunas micras hasta algunos centímetros.
Existen no obstante ejemplos de células
visibles a simple vista: como el huevo de las
aves, cuyo volumen está determinado por
la enorme acumulación de materiales de
reserva. Las dimensiones de las células no
varían con las del organismo del que forman
parte; por ejemplo, el volumen de las
células de la mucosa intestinal del
ratón no difiere mucho del de las
células análogas del elefante.
Constituyen una excepción a esta regla los
elementos llamados perennes, como las
células nerviosas y musculares.
(2)
-
Antiguamente los biólogos
pensaban que las células estaban
formadas por una gelatina uniforme que llamaban
protoplasma. Con la microscopía
electrónica y otras herramientas modernas de
investigación, se ha
extendido la percepción del mundo con
respecto a las células. En la actualidad
sabemos que la célula tiene un alto
nivel de organización y que es
sorprendentemente compleja: tienen su propio
centro de control, su sistema de transporte interno, fuentes de energía,
fábricas para procesar la materia que
requiere, plantas de empaquetamiento, e
incluso un sistema de autodestrucción.
En nuestros días el término
protoplasma si acaso se utiliza es en un
sentido muy general. La porción de
protoplasma que se encuentra fuera del
núcleo se llama citoplasma, y el
material interno del núcleo se llama
nucleoplasma. Los organelos se encuentran
suspendidos en el componente líquido del
citoplasma y del nucleoplasma. Cada uno de los
organelos delimitados por sus membranas forma
uno o más compartimentos independientes
dentro del citoplasma. (3)
Estructura de las células
eucariotas y sus funciones: (4)
Cuadro 2.2.2-1.
Estructura de la célula eucariota y sus
funciones
|
Estructura
|
Descripción
|
Función
|
Núcleo
celular
|
|
|
Núcleo
|
Gran estructura rodeada por una
doble membrana; contiene nucleolo y
cromosomas.
|
Control de la
célula
|
Nucleolo
|
Cuerpo granular dentro del
núcleo; consta de ARN y proteínas.
|
Lugar de síntesis
ribosómica; ensamble de
subunidades
ribosómicas.
|
Cromosomas
|
Compuestos de un complejo de
ADN y proteínas,
llamado cromatina; se observa en forma de
estructuras en cilindro durante la
división celular.
|
Contiene genes (unidades de
información
hereditaria que gobiernan la estructura y
actividad celular).
|
Sistema de membranas de la
célula.
|
Membrana celular (membrana
plasmática)
|
Membrana limitante de la
célula viva
|
Contiene al citoplasma; regula
el paso de materiales hacia dentro y
fuera de la célula; ayuda a
mantener la forma celular; comunica a la
célula con otras.
|
Retículo
endoplasmático (ER)
|
Red de membranas internas que se
extienden a través del
citoplasma.
|
Sitio de síntesis de
lípidos y de
proteínas de membrana; origen de
vesículas intracelulares de
transporte, que acarrean proteínas
en proceso de
secreción.
|
Liso
|
Carece de ribosomas en su
superficie externa.
|
Biosíntesis de
lípidos; Destoxicación de
medicamentos.
|
Rugoso
|
Los ribosomas tapizan su
superficie externa.
|
Fabricación de muchas
proteínas destinadas a
secreción o incorporación
en membranas.
|
Ribosomas
|
Gránulos compuestos de
ARN y proteínas; algunos unidos al
ER, otros libres en el
citoplasma.
|
Síntesis de
polipéptidos.
|
Aparato de Golgi
|
Compuesto de saculaciones
membranosas planas.
|
Modifica, empaca (para
secreción) y distribuye
proteínas a vacuolas y a otros
organelos.
|
Lisosomas
|
Sacos membranosos (en
animales).
|
Contienen enzimas que degradan
material ingerido, las secreciones y
desperdicios celulares.
|
Vacuolas
|
Sacos membranosos (sobre todo en
plantas, hongos y algas )
|
Transporta y almacena material
ingerido, desperdicios y agua.
|
Microcuerpos (ej.
peroximas)
|
Sacos membranosos que contienen
una gran diversidad de
enzimas.
|
Sitio de muchas reacciones
metabólicas del
organismo.
|
Organismos transductores de
energía
|
Mitocondrias
|
Sacos que constan de dos
membranas; la mambrana interna
está plegada en
crestas.
|
Lugar de la mayor parte de las
reacciones de la respiración
celular; transformación en ATP, de
la energía proveniente de la
glucosa o lípidos.
|
Plástidos
|
Sistema de tres membranas: los
cloroplastos contienen clorofila en las
membranas tilacoideas
internas.
|
La clorofila captura
energía luminosa; se producen ATP
y otros compuestos energéticos,
que después se utilizan en la
conversión de CO2 en
glucosa.
|
Citoesqueleto
|
Microtúbulos
|
Tubos huecos formados por
subunidades de tubulina.
|
Proporcionan soporte
estructural; intervienen en el movimiento y
división celulares; forman parte
de los cilios, flagelos y
centriolos.
|
Microfilamentos
|
Estructuras sólidas,
cilíndricas formadas por
actina.
|
Proporcionan soporte
estructural; participan en el movimiento
de las células y organelos,
así como en la división
celular.
|
Centriolos
|
Par de cilindros huecos cerca
del centro de la célula; cada
centriolo consta de 9 grupos de 3
microtúbulos.
|
Durante la división
celular en animales se forma un uso
mitótico entre ambos centriolos;
en animales puede iniciar y organizar la
formación de microtúbulos;
no existen en las plantas
superiores.
|
Cilios
|
Proyecciones más o menos
cortas que se extienden de la superficie
celular; cubiertos por la membrana
plasmática; compuestos de 2
microtúbulos centrales y 9 pares
periféricos
|
Locomoción de algunos
organismos unicelulares; desplazamiento
de materiales en la superficie celular de
algunos tejidos.
|
Flagelos
|
Proyecciones largas formadas por
2 microtúbulos centrales y 9
periféricos; se
extienden desde la superficie celular;
recubiertos por mambrana
plasmática.
|
Locomoción de las
células espermáticas y de
algunos organismos
unicelulares.
|
-
ORGANIZACIÓN
UNICELULAR:
Células
Eucariotas:
Los organismos Eucariotas son
aquellos que contienen una estructura
llamada núcleo, que se encuentra
limitado por una membrana. El núcleo
sirve para localizar el material
genético, el ADN.
El término eucariota
significa "núcleo verdadero" y se
refiere a que el material genético
de las células, está incluido
en un núcleo distinto, rodeado por
una membrana nuclear. Estas células
también presentan varios organelos
limitados por membranas que dividen el
citoplasma celular en varios compartimentos
adicionales. Algunos organelos sólo
se presentan en algunas variedades
celulares específicas. Por ejemplo,
los cloroplastos, que atrapan la luz solar para
conversión de energía, se
hallan en las células que realizan
fotosíntesis. Los
organelos especializados de las
células eucariotas les permiten
resolver algunos de los problemas relacionados con
su gran tamaño, de manera que pueden
ser considerablemente más grandes
que las células procariotas.
(5)

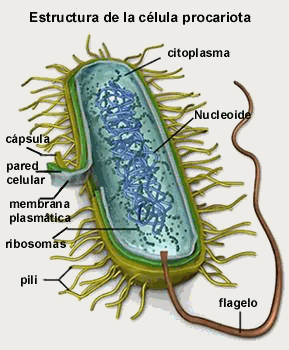
Células
procariotas:
Las células procariotas son
aquellas que carecen de núcleo,
vacuolas, mitocondrias y otros
orgánulos subcelulares, generalmente
son más pequeñas que las
eucariotas. Son organismos de una sola
célula que pertenecen al grupo
Monera: se incluyen bacterias y algas
verdeazules o cianobacterias, que no son
sino bacterias fotosintéticas. El
ADN de las células procariotas
está confinado a una o más
regiones nucleares, que a veces se
denominan nucleoides, los cuales no
están limitados por una membrana
independiente.
Las células procariotas
tienen una membrana plasmática que
confina el contenido celular a un
compartimento interno, pero carece de un
sistema de membranas internas en forma de
organelos. En algunas células
procariotas la membrana plasmática
puede plegarse hacia adentro y forma un
complejo de membranas internas en donde se
piensa se llevan a cabo las reacciones de
transformación de energía.
Algunas células procariotas
también tienen una pared celular o
membrana externa, que es una estructura que
encierra a toda la célula, incluida
la membrana plasmática.
(6)
Virus:
Los virus o viriones no son seres
celulares, dado a que no se mueven por si
mismos y no son capaces de metabolizar de
manera independiente: sólo pueden
vivir cuando han infectado una
célula. Un virus consiste en un
filamento de ADN o de ARN (pero nunca ambos
ácidos nucleicos en
un mismo virus) contenido en una envoltura
proteica de forma geométrica
denominada cápside o cápsida,
que está integrada por un conjunto
de subunidades idénticas, los
capsómeros, dispuestas en mosaico.
Los virus pueden presentarse desnudos o
bien revestidos de una envoltura
lipoproteica, procedente de una
porción de la membrana
plasmática de la célula
huésped. (7)

ORGANIZACIÓN
PLURICELULAR: (8)
Tejidos
vegetales:
Los vegetales presentan dos
tipos de organización
celular bien diferenciados. Los hongos, las
algas y las plantas no vasculares en
general carecen de verdaderos tejidos y
vasos; las divisiones celulares tienen
lugar en un plano y los nutrientes llegan
directamente a todas las células sin
necesidad de un sistema especializado de
transporte. Es la organización tipo
talo.
Las plantas vasculares, adaptadas
ya a la vida terrestre y aérea, son
las que tienen verdaderos tejidos
diferenciados. Es la organización
tipo cormo, con raíces, hojas
y tallos verdaderos, los tres
órganos de las plantas
superiores.
Tejidos
animales:
Generalmente los tejidos animales
se clasifican en cuatro tipos
fundamentales: epitelial, conectivo,
muscular y nervioso. Otras clases de
tejidos como el óseo o la sangre son formas de tejido
conectivo.
Los tejidos se reúnen en
forma de órganos especializados en
un tipo de actividad (corazón,
estómago, etc.), de forma que un
determinado órgano puede estar
formado por varios tejidos diferentes. Un
conjunto de órganos coordinados
puede desarrollar una función
específica (por ejemplo, la
respiración) constituye un aparato.
Y un conjunto de órganos de la misma
clase de tejidos y distribuidos por todo el
organismo con una misma función es
lo que se llama un sistema (sistema nervioso, endocrino,
etc.).
- PROCESOS CELULARES:
(9)
-
Metabolismo:
En todos los seres vivos
ocurren reacciones
químicas esenciales para la
nutrición, el
crecimiento y la reparación de
las células, así como
para la conversión de la
energía en formas utilizables.
La suma de todas estas actividades
químicas del organismo recibe el
nombre de metabolismo. Las
reacciones metabólicas ocurren
de manera continúa en todo ser
vivo; en el momento en que se suspenden
se considera que el organismo ha
muerto.
Crecimiento:
Algunas cosas no vivas parecen
crecer. Por ejemplo, se forman
cristales en una solución
sobresaturada de una sal; a medida que
va saliendo más sal de la
solución, los cristales crecen
más y más. No obstante,
ese proceso no es crecimiento en el
sentido biológico. Los
biólogos restringen el
término crecimiento a los
procesos que incrementan la cantidad de
sustancia viva en el organismo. El
crecimiento por tanto es, es un aumento
de la masa celular, como resultado de
un incremento del tamaño de las
células individuales del
número de células, o de
ambos. El crecimiento puede ser
uniforme en las diversas partes del
organismo, o mayor en unas partes que
en otras, de modo que las proporciones
corporales cambian conforme ocurre el
crecimiento.
Algunos organismos p. ej. casi
todos los árboles siguen
creciendo en forma definida. Muchos
animales tienen un periodo de
crecimiento, el cual termina cuando se
alcanza el tamaño característico
del adulto. Uno de los aspectos
más notables del proceso es que
cada parte del organismo sigue
funcionando conforma éste
crece.
Movimiento:
El movimiento, aunque no
necesariamente la locomoción, es
otra característica de los seres
vivos. El movimiento de casi todos los
animales es muy obvio: se agitan,
raptan, nadan, corren o vuelan. Los
movimientos de las plantas son mucho
más lentos y menos obvios, pero
no por ello dejan de ser un hecho. El
movimiento de flujo del material vivo
en el interior de las células de
las hojas de las plantas se conoce como
ciclosis.
La locomoción puede ser
el resultado de la actividad de
diminutas extensiones piliformes
llamadas cilios o flagelos, de la
contracción de los
músculos, o del lento flujo de
una masa de sustancias celulares
llamado movimiento
amiboideo.
Irritabilidad:
Los seres vivos reaccionan a
los estímulos, que son cambios
físicos o químicos en su
ambiente interno o externo. Los
estímulos que evocan una
reacción en la mayoría de
los organismos son: cambios de color, intensidad o
dirección de la
luz; cambios en temperatura, presión o
sonido, y cambios en la
composición química del
suelo, aire o agua
circundantes. En los animales
complejos, como el ser humano, ciertas
células del cuerpo están
altamente especializadas para
reaccionar a ciertos tipos de
estímulos; por ejemplo las
células de la retina del ojo
reaccionan a la luz. En los organismos
más simples esas células
pueden estar ausentes, pero el
organismo entero reacciona al
estímulo. Ciertos organismos
celulares reaccionan a la luz intensa
huyendo de ella.
La irritabilidad de las
plantas no es tan obvia como la de los
animales, pero también los
vegetales reaccionan a la luz, la
gravedad, el agua y otros
estímulos, principalmente por
crecimiento de su cuerpo. El movimiento
de flujo del citoplasma de las
células vegetales se acelera o
detiene a causa de las variaciones en
la intensidad de la luz.
Reproducción:
Aunque hubo una época
en la que se creía que los
gusanos se creaban a partir de crines
de caballo sumergidas en abrevaderos,
que los gusanos de la carne se
originaban a partir de ésta, y
que las ranas surgían del fango
del Nilo, ahora se sabe que cada uno de
esos organismos sólo puede
provenir de organismos preexistentes.
Uno de los principios fundamentales
de la biología es que "toda vida
proviene exclusivamente de los seres
vivos". Si existe alguna
característica que pueda
considerarse la esencia misma de la
vida, ésta es la capacidad que
tienen los organismos de
reproducirse.
En el caso de los organismos
más simples, como las amibas, la
reproducción puede ser asexual;
es decir, sin contacto sexual. Cuando
una amiba alcanza cierto tamaño,
se reproduce partiéndose en dos,
y forma dos amibas nuevas. Antes de
dividirse, cada amiba produce un
duplicado de su material
genético, de modo que cada amiba
hija, es idéntica a la
célula progenitora.
En casi todas las plantas y
animales, la reproducción sexual
se realiza mediante la
reproducción de células
especializadas llamadas óvulos y
espermatozoides, las cuales se unen y
forman el óvulo fecundado, o
cigoto, del que nace el nuevo
organismo. Cuando la
reproducción es sexual, cada
descendiente es el producto de la
interacción de diversos genes,
aportados por la madre y el padre, en
vez de ser idéntico al
progenitor, como sucede en el proceso
asexual. La variación genética es la
materia prima sobre la
cual actúan los procesos vitales
de la evolución y la
adaptación.
Adaptación:
La capacidad que muestra una especie para
adaptarse a su ambiente es la
característica que les permite
sobrevivir en un mundo en constante
cambio. Las adaptaciones
son rasgos que incrementan la capacidad
de sobrevivir en un ambiente
determinado. Dichas adaptaciones pueden
ser estructurales, fisiológicas
o conductuales, o una
combinación de ellas.
La adaptación trae
consigo cambios en la especie, mas que
en el individuo. Si todo organismo de
una especie fuera exactamente
idéntico a los demás,
cualquier cambio en el ambiente
sería desastroso para todos
ellos, de modo que la especie se
extinguiría. La mayor parte de
las adaptaciones se producen durante
periodos muy prolongados de tiempo, y en ellas
intervienen varias generaciones. Las
adaptaciones son resultado de los
procesos evolutivos.
-
HOMEOSTASIS:
En todos los organismos,
los diversos procesos
metabólicos deben ser
cuidadosos y constantemente
regulados para mantener un estado de
equilibrio.
Cuando ya se sintetizó una
cantidad suficiente de un
componente celular, es necesario
reducir su producción
o suspenderla por completo. Cuando
declina la cantidad de
energía disponible en una
célula, es necesario que
entren en funcionamiento los
procesos adecuados para poner a
disposición de la
célula nueva energía.
Estos mecanismos autorregulados de
control son notablemente sensibles
y eficientes. La tendencia de los
organismos a mantener un medio
interno constante se denomina
homeostasis, y
los mecanismos que realizan esa
tarea se llaman mecanismos
homeostáticos.
La regulación de la
temperatura corporal en el ser
humano es un ejemplo de la
operación de tales
mecanismos. Cuando la temperatura
del cuerpo se eleva por arriba de
su nivel normal de 37°C, la
temperatura de la sangre es
detectada por células
especializadas del cerebro que
funcionan como un termostato.
Dichas células envían
impulsos nerviosos hacia las
glándulas sudoríparas
e incrementan la secreción
del sudor. La evaporación
del sudor que humedece la
superficie del cuerpo reduce la
temperatura corporal. Otros
impulsos nerviosos provocan la
dilatación de los capilares
sanguíneos de la piel, haciendo
que esta se sonroje. El aumento de
flujo sanguíneo en la piel
lleva más calor hacia la
superficie corporal para que desde
ahí se disipe la radiación.
Cuando la temperatura del
cuerpo desciende por debajo de su
nivel normal, el sensor del cerebro
inicia una serie de impulsos que
constriñen los vasos
sanguíneos de la piel,
reduciendo así la
pérdida de calor a
través de la superficie. Si
la temperatura corporal desciende
aún más, el cerebro
empieza a enviar impulsos nerviosos
hasta los músculos,
estimulando las rápidas
contracciones musculares conocidas
como escalofríos, un proceso
que tiene como resultado la
generación de calor.
(10)
TRANSPORTE A TRAVES DE
MEMBRANA: (11)
El hecho de que una
membrana permita el paso de las
moléculas de cierta
sustancia depende de la estructura
de aquella y el tamaño y
carga eléctrica de las
moléculas. Se dice que una
membrana es permeable para alguna
sustancia si permite que
ésta la cruce e impermeable
si no permite el paso de dicha
sustancia. Una membrana
selectivamente permeable permite el
paso de algunas sustancias pero no
el de otras. Todas las membranas
biológicas que rodean las
células, núcleos,
vacuolas, mitocondrias,
cloroplastos y otros organelos
celulares son selectivamente
permeables.
Al reaccionar a las
condiciones ambientales cambiantes
a las diversas necesidades de la
célula, la membrana puede
constituir una barrera al paso de
un compuesto determinado en cierto
momento, mientras promueve
activamente su paso en otro
momento. Mediante la
regulación del
tráfico químico de
esa manera, la célula
controla su propia
composición interna de iones
y moléculas que puede ser
muy diferente a la del exterior. En
el mundo abiótico, los
materiales se mueven pasivamente
por procesos físicos como la
difusión. En los seres
bióticos, los materiales
también se mueven
activamente por procesos
fisiológicos como transporte
activo, exocitosis y endocitosis.
Esos procesos fisiológicos
activos
demandan un gasto de energía
por parte de la
célula.
Difusión:
Algunas sustancias se
desplazan hacia adentro y afuera de
las células, y se mueven
dentro de éstas por medio de
un proceso llamado difusión
simple, el cual se basa en el
desplazamiento al azar. A
temperaturas mayores al cero
absoluto, todos los átomos y
moléculas poseen
energía cinética, o
energía de movimiento. Los
tres estados de la materia
(sólido, liquido y gaseoso)
difieren con respecto a la libertad del
movimiento de las moléculas
que los constituyen. Las
moléculas del estado
sólido se encuentran muy
cerca una de otra, y las fuerzas de
atracción entre ellas les
permiten vibrar, pero no
desplazarse. Las moléculas
de un líquido se encuentran
más separadas entre
sí, respecto a las de un
sólido; las fuerzas
intermoleculares son más
débiles, y las
moléculas se desplazan con
relativa libertad; en el estado
gaseoso, las moléculas
están tan separadas que las
fuerzas intermoleculares son
mínimas y por tanto el
desplazamiento de las
moléculas sólo
está limitado por las
paredes del recipiente que las
contiene. Esto significa que los
átomos y moléculas de
líquidos y gases se
separan en una especie de
"desplazamiento al azar". A este
desplazamiento se atribuye el
proceso de difusión,
movimiento neto de
partículas (átomos,
iones, moléculas, etc.) de
una región de alta
concentración a una de
concentración más
baja, de manera que las
partículas adquieren una
distribución
uniforme. Por tanto puede decirse
que la difusión implica el
movimiento neto de
partículas a favor de un
gradiente de concentración.
Esto no significa que las
partículas no puedan
desplazarse en sentido contrario al
gradiente de concentración.
Sin embargo, si al inicio hay mayor
cantidad de partículas en
una región de
concentración elevada,
lógicamente más
partículas se
desplazarán desde la zona de
alta concentración hacia la
de baja concentración, que a
la inversa. La proporción de
difusión está en
función del tamaño y
forma de las moléculas, de
sus cargas eléctricas y de
la temperatura, las
moléculas se mueven con
mayor rapidez y aumenta la
proporción de
difusión.
Diálisis:
La difusión de un
soluto a través de una
membrana diferencialmente permeable
se llama diálisis. Para
demostrar la diálisis se
utiliza una bolsa de celofán
llena con una solución de
azúcar,
que luego se sumerge en un matraz
que contiene agua pura. Si la
membrana de celofán es
permeable al azúcar y al
agua, las moléculas de
azúcar pasarán a
través de ella hasta que la
concentración de
azúcar en el agua de los dos
lados de la membrana sea
exactamente igual. A partir de ese
momento, las moléculas de
soluto (así como
también las moléculas
de agua) seguirán pasando a
través de la membrana, pero
ya no habrá ningún
cambio neto en las concentraciones
ya que la velocidad de
movimiento será igual en
ambos sentidos. La diálisis
renal es una aplicación
práctica de este proceso;
los productos de
desecho, que se difunden a
través de las membranas
artificiales del aparato, pueden
retirarse del organismo, pero los
eritrocitos, proteínas
sanguíneas y otras
moléculas grandes, no se
difunden a través de la
membrana, y por tanto se
retendrán en el
organismo.
Osmosis:
La ósmosis es una
variedad especial de
difusión que implica el
movimiento de moléculas
solventes (p. ej., agua) a
través de una membrana de
permeabilidad selectiva. Las
moléculas de agua pasan
libremente en cualquier
dirección, pero al igual que
en todos los procesos de
difusión, el movimiento neto
ocurre a partir de la región
de mayor concentración a la
de menor. La mayor parte de los
solutos no puede difundirse
libremente a través de la
membrana celular de permeabilidad
selectiva.
Los principios que
intervienen en el proceso de
ósmosis se ilustran mediante
la utilización de un aparato
llamado tubo en U, éste se
divide en dos secciones por una
membrana de permeabilidad selectiva
que permite que las
moléculas de soluto
(glucosa, sal y otras). En una
parte del tubo se coloca una
solución de agua y solutos;
en la otra se coloca agua pura. La
solución de agua y solutos
contiene una concentración
de agua menor a la del agua pura,
porque las moléculas de
soluto han diluido las
moléculas de agua. Por
tanto, hay un movimiento neto de
moléculas de agua del lado
del agua pura (con mayor
concentración de
moléculas de agua) hacia el
lado del agua con soluto (que tiene
menor concentración de
moléculas de agua), como
resultado de esto el nivel del
líquido del lado del agua
pura disminuye, mientras que se
eleva el del lado del agua con
soluto. Sin embargo aún
existe una diferencia en la
concentración de las
moléculas de agua entre
ambos lados, debido a que las
moléculas de soluto no
pueden moverse a través de
la membrana. El movimiento neto de
agua continuará, y el nivel
del líquido del lado del
agua con soluto seguirá
aumentando. En condiciones no
sujetas a la gravedad, este proceso
continuará indefinidamente,
pero en la tierra, el
peso de la columna del
líquido en aumento
finalmente ejercerá una
presión suficiente para
detener el cambio en los niveles
del líquido, aunque las
moléculas de agua
continuarán pasando a
través de la membrana en
ambas direcciones.
La presión
osmótica de una
solución se define como la
tendencia que presenta el agua de
moverse hacia dicha solución
mediante ósmosis.
Soluciones
isotónicas,
hipertónicas e
hipotónicas:
Con frecuencia deseamos
comparar las presiones
osmóticas de dos soluciones. En
todo líquido de los
compartimentos de las
células vivas se encuentran
disueltas sales, azúcares y
otras sustancias que le confieren a
dicho líquido una
determinada presión
osmótica. Cuando una
célula se coloca en una
solución, cuya
presión osmótica es
igual a la suya, no hay movimiento
neto de partículas de agua,
ni hacia fuera ni hacia dentro de
ella; por tanto la célula no
se hincha ni se encoge. Se dice que
el líquido en el cual se
colocó la célula es
un líquido
isotónico (es decir
que tiene presión
osmótica igual) con respecto
al líquido interior de la
célula.
Si el líquido
circundante tiene una
concentración de solutos
mayor que la del líquido
intracelular y, por tanto, una
presión osmótica
mayor que la de éste se dice
que es una solución
hipertónica; una
célula colocada en una
solución hipertónica
pierde agua y por tanto, se encoge.
Cuando una célula con pared
celular se coloca se coloca en un
medio hipertónico pierde
agua, y entonces su contenido
disminuye dentro de la pared
celular; este proceso se llama
plasmólisis.
Si el líquido
circundante posee una
concentración de solutos
menor que la del líquido
intacelular, y por tanto tiene una
presión osmótica
menor que la de éste, se
denomina solución
hipotónica; en estas
circunstancias, el líquido
se desplazará hacia el
interior de la célula
provocando que ésta se
hinche.
Presión de
turgencia:
Las paredes celulares
rígidas de células
vegetales, algas, bacterias y
hongos hacen posible que esos
organismos vivan sin reventar en un
medio externo muy diluido, que
contenga una concentración
muy baja de solutos. Debido a las
sustancias disueltas en el
citoplasma, las células son
hipertónicas respecto al
medio externo. (El medio
circundante es hipotónico
respecto al citoplasma). El agua
tiende a difundirse hacia el
interior de las células por
ósmosis, llenando sus
vacuolas centrales y
distendiéndolas. La
célula se hincha acumulando
presión, llamada
presión de turgencia, contra
las paredes celulares
rígidas de celulosa. La
pared celular puede estirarse muy
poco, y se alcanza un estado de
equilibrio cuando su resistencia
impide que la célula se
hinche más. Al llegar a este
punto ya no hay movimiento neto de
moléculas de agua hacia el
interior de la célula
(aunque desde luego, las
moléculas continúan
moviéndose hacia dentro y
hacia fuera de la membrana). La
presión de turgencia es un
factor importante en el
sostén del cuerpo de las
plantas herbáceas. Por este
motivo, una flor se marchita cuando
la presión de turgencia de
sus células disminuye (las
células han sufrido
plasmólisis) por falta de
agua.
Transporte mediado de
moléculas
pequeñas:
La membrana celular es
relativamente impermeable a casi
todas las grandes moléculas
polares. Esto constituye una
ventaja biológica para la
célula, ya que casi todos
los compuestos metabolizados en su
interior son polares y la
impermeabilidad de la membrana
impide su pérdida por
difusión. Para transportar
nutrientes polares, como glucosa y
aminoácidos, a través
de la membrana lipídica
hacia el interior de la
célula, han aparecido por
evolución sistemas de
proteínas transportadoras
que se unen a esas moléculas
y las transfieren a través
de la membrana. El paso de solutos
a través de la membrana
celular por el sistema de
transporte se llama transporte
mediado. La energía que se
necesita proviene de dos fuentes:
difusión facilitada y
transporte activo.
Difusión
facilitada:
En los casos más
simples, la célula utiliza
la energía almacenada por el
gradiente de concentración
de una sustancia cuya
concentración es mayor en el
líquido extracelular que el
intracelular. En estas
circunstancias, mientras la
membrana sea permeable a dicha
sustancia, ésta se
desplazará hacia el interior
de la célula. Este tipo de
transporte se llama difusión
facilitada. La difusión
facilitada depende de la existencia
de proteínas
transportadoras, las cuales se
combinan temporalmente con la
molécula de soluto para
acelerar el paso de ésta a
través de la membrana
celular. La proteína
transportadora no se modifica por
ésta acción;
después de transportar la
molécula de soluto, queda
libre para unirse a una nueva
molécula.
Transporte activo
mediado:
Algunas moléculas
se transportan a través de
la célula mediante el
proceso de difusión; otras
las requiere la célula en
concentraciones mayores a su
concentración extracelular.
Estas moléculas se
incorporan mediante mecanismos de
transporte activo. Este mecanismo
exige una fuente de energía
debido a que el transporte activo
implica el "bombeo" de una
molécula en contra de su
gradiente de concentración.
Por tanto los sistemas de
transporte activo utilizan
energía generada por el
metabolismo celular en forma de
trifosfato de adenosina (ATP) o
bien utilizan algún otro
tipo de energía almacenada,
derivada de la hidrólisis
del ATP.
Uno de los ejemplos
más sorprendentes de los
mecanismos de transporte activo es
la bomba de sodio y potasio que se
observa en todas las células
animales.
Sistema de
cotransporte:
El gradiente
electroquímico generado por
la bomba de sodio y potasio
también proporciona
suficiente energía para
propiciar el transporte activo de
otras moléculas esenciales.
En estas reacciones, el gradiente
de concentración de sodio y
potasio cotransporta las
moléculas requeridas, junto
con los iones de sodio y potasio.
La energía del ATP se usa en
forma indirecta al favorecer el
transporte activo de una
molécula requerida, cuando
une el transporte de dicha
molécula en contra de su
gradiente de concentración,
con el transporte de sodio o
potasio, a favor de su gradiente de
concentración.
Sistema de transporte
múltiple
integrado:
En algunas células
se observa el funcionamiento de
más de un sistema de
transporte para una sustancia
determinada. Por ejemplo, el
transporte de la glucosa del
intestino hacia el torrente
circulatorio se lleva a cabo a
través de una delgada capa
de células epiteliales que
recubren la luz del intestino y que
poseen regiones especializadas, o
dominios, en su membrana
plasmática. La superficie de
éstas células,
expuesta en el intestino, posee una
gran cantidad de microvellosidades
que incrementan con eficacia la
superficie de membrana disponible
para absorción. El
transporte de glucosa en esta zona
de la superficie celular es parte
de un sistema de transporte activo
que se efectúa en
cotransporte con el sodio. La
concentración intracelular
de sodio se mantiene en cifras
bajas por función de una
bomba de sodio y potasio en la
superficie opuesta de la
célula, que bombea el sodio
hacia el torrente circulatorio.
Gracias a su elevada
concentración dentro de la
célula, la glucosa puede ser
transportada hacia el torrente
circulatorio mediante
difusión
facilitada.
Transporte de grandes
moléculas a través de
las membranas:
En la difusión
simple, en la difusión
facilitada y en el transporte
activo las moléculas
individuales y los iones pasan a
través de la membrana
celular. Sin embargo en ocasiones
también es necesario el
desplazamiento de cantidades
más grandes de material o de
partículas de alimento o
incluso de células
completas, hacia afuera o adentro
de una célula. Esto implica
un gasto de energía por
parte de la célula y en
ocasiones conlleva también
la fusión
de membranas. En la exocitosis una
célula expulsa productos de
desecho o productos
específicos de
secreción (como hormonas),
mediante la fusión de una
vesícula con la membrana
plasmática de la
célula. La exocitosis
consiste en la fusión de la
membrana de la vesícula
secretora con la membrana
plasmática.
En la endocitosis, la
célula incorpora materiales
hacia su interior. En los sistemas
biológicos operan varios
mecanismos endocitóticos.
Por ejemplo en la fagocitosis, la
célula ingiere
partículas sólidas
como bacterias o nutrientes.
Durante la ingestión los
pliegues de la membrana celular
engloban a la partícula, que
se ha unido a la superficie
celular, y forman una vacuola
alrededor de ella. Una vez que la
membrana ha encerrado a la
partícula en
cuestión, se4 fusiona en el
punto de contacto, dejando que la
vacuola flote libremente en el
citoplasma. Posteriormente la
vacuola se fusiona con los
lisosomas, donde el material es
ingerido y degradado.
En otro tipo de
endocitosis llamada pinocitosis, la
célula incorpora materiales
disueltos. Algunos pliegues de la
membrana plasmática engloban
gotas de líquido, las cuales
emergen en el citoplasma en forma
de pequeñas
vesículas.
Otro tipo de endocitosis
llamado endocitosis mediada por
receptor, algunas proteínas
específicas de determinadas
partículas se unen a
proteínas receptoras,
localizadas en la membrana
plasmática de la
célula. Luego, las
moléculas ligadas al
receptor emigran en placas
recubiertas, que son regiones de la
superficie citoplasmática de
la membrana recubiertas con
estructuras en forma de
cepillo.
- FOTOSINTESIS Y
RESPIRACION:
- HOMEOSTASIS Y TRANSPORTE
A TRAVES DE MEMBRANA:
- TIPOS DE ORGANIZACIÓN
CELULAR:
- ESTRUCTURA CELULAR
GENERAL:
- LA CELULA:
- ORGANIZACIÓN Y PROCESOS:
- INTRODUCCION:
FOTOSINTESIS: (12)
La fotosíntesis es una de las funciones
biológicas fundamentales. Por medio de la clorofila
contenida en los cloroplastos, los vegetales verdes son capaces
de absorber la energía que la luz solar emite como
fotones y transformarla en energía química. Esta
se acumula en las uniones químicas producidas por la
síntesis de muchos principios nutritivos. Las
mitocondrias son susceptibles de utilizar y transformar la
energía contenida en las sustancias alimenticias
mediante la fosforilación oxidativa. En la
fotosíntesis ocurre, hasta cierto punto, un proceso
inverso. En los procariotas fotosintéticos, bacterias y
cianofíceas, no hay cloroplastos, pero la clorofila se
encuentra en extensos sistemas membranosos internos.
Los cloroplastos y las mitocondrias tienen muchas
semejanzas estructurales y funcionales, pero también
poseen algunas diferencias.
La principal reacción de la fotosíntesis
es:
Luz, clorofila
nCO2 + H2O (CH2O)n
+ nO2 (1)
que consiste en la combinación de
CO2 y H2O para formar carbohidratos con liberación de
O2.
Se ha calculado que cada molécula de
CO2 de la atmósfera, se incorpora al vegetal cada
200 años, y que el oxígeno del aire es renovado por las
plantas cada 2000 años. Sin plantas no existiría
O2 en la atmósfera y la vida sería
casi imposible.
Los carbohidratos formados al principio por la
fotosíntesis son azúcares solubles que pueden
acumularse como granos de almidón o de otros
polisacáridos dentro del cloroplasto o, más
frecuentemente, en el interior de los leucoplastos
(amiloplastos). Después de varias etapas que involucran
la participación de diferentes tipos de plástidos
y de sistemas enzimáticos, el material
fotosintético se almacena como un producto de reserva o
bien se emplea como una parte estructural del vegetal (ej.
celulosa).
Desde los primeros estudios, se sugirió que en
la reacción (1) el H2O era el dador de
hidrógeno, del mismo modo que el H2S es el dador en las
sulfobacterias.
luz
2H2S + CO2 (CH2O) +
2S + H2O (2)
Por tanto, la reacción (1) en las plantas
superiores puede expresarse:
luz
2nH2O + nCO2 (CH2O)n
+ nH2O + nO2 (3)
La reacción (3) demuestra que el agua es el
dador de H2 y que todo el O2 liberado
proviene de ella.
Los estudios bioquímicos revelaron que la
reacción (3) estaba compuesta por una serie de pasos
complejos, de los cuales algunos se producen sólo en
presencia de luz, mientras que los otros pueden llevarse a cabo
también en la oscuridad. Por lo tanto, se las denomina
reacciones en la luz y en la oscuridad. En la primera, la luz
es absorbida y empleada por la clorofila; ésta es la
reacción fotoquímica o de Hill. En la segunda,
tiene lugar la fijación y reducción del
CO2 por medio de mecanismos
termoquímicos.
 Reacción fotoquímica:
Reacción fotoquímica:
Al estudiar la reacción fotoquímica es
necesario tener presente el proceso de la oxidación
fosforilativa de las mitocondrias. En ésta el flujo de
los electrones va desde NADH2 hacia O2, siendo el
gradiente de potencial de oxidoreducción (-0,6 a
+0,81V). En la fotosíntesis es a la inversa: los
electrones fluyen desde el H2O al NADPH2,
es decir de +0,81 a –0,6 V.
La reacción fotoquímica o primaria de la
fotosíntesis tiene lugar en las membranas de los
tilacoides. Cuando éstas son iluminadas se produce la
transferencia de electrones desde el agua (E'o=0,81V) al
aceptor final (E'o=-0,6V). Para que esa transferencia pueda
cumplirse en contra del gradiente electroquímico es
indispensable la energía provista por los fotones de
luz. Este proceso tiene lugar en una cadena de transportadores
de electrones acoplados a la fosforilación del ADP a
ATP.
A diferencia de la fosforilación oxidativa de
las mitocondrias, en la fotofosforilación no se emplea
O2. Mediante la fotofosforilación, los
vegetales verdes pueden producir una cantidad de ATP 30 veces
mayor que la obtenida en sus propias mitocondrias. Por otra
parte éstas plantas contienen muchos más
cloroplastos que mitocondrias.
Ciclo fotosintético de la reducción
del carbono o
ciclo de Calvin:
Las moléculas de ATP y NADPH2,
producidas en el tilacoide proveen, a las enzimas del estroma
del cloroplasto, de la energía necesaria para fijar el
CO2 y sintetizar los carbohidratos. Junto con la
energía proporcionada por el ATP, el NADPH2
reducido, es capaz de reducir el CO2
atmosférico y combinarlo con el hidrógeno para
formar los diferentes carbohidratos. Este proceso comprende
diversos pasos que han sido dilucidados fundamentalmente
mediante el uso de CO2 radiactivo. Las reacciones
involucradas son tan rápidas que se completan en un
segundo o menos después del agregado de
C14O2. Tienen lugar en la oscuridad
total, si las plantas fueron expuestas previamente a la
luz.
En células expuestas al
C14O2 durante 5 segundos, el compuesto
predominante es el 3-fosfoglicerato. Dos moléculas de
triosafosfato se unen para formar hexosa (fructosa) difosfato,
de la que deriva luego la glucosa fosfato. A partir de
ésta última se originan varios disacáridos
y polisacáridos.
La enzima inicial del ciclo de Calvin es la ribulosa
1,5 difosfato carboxilasa o carboxidismutasa, que tiene un alto
peso molecular (500000 dalton), con ocho subunidades grandes
producidas en el cloroplasto y ocho más pequeñas
sintetizadas en el citosol. Esta enzima representa la mitad de
las proteínas del estroma y desempeña el papel
central del ciclo al combinar una pentosa-ribulosa 1,5
difosfato con un CO2 para producir dos
moléculas de 3-fosfoglicerato, utilizando una de
H2O.
Esta triosa se fosforila luego con ATP y se forma una
molécula activada que puede aceptar H+ y
electrones a partir de NADPH. Se trata del
3-fosfogliceraldehido, que luego se reduce para constituir
hexosas y carbohidratos más complejos.
Calvin y Benson propusieron que en cada vuelta de
éste ciclo (también llamado ciclo C3) se regenera
una molécula de ribulosa 1,5 difosfato. Tal proceso de
regeneración es muy complejo y comprende unas 12
reacciones enzimáticas. Para producir una hexosa a
partir de la fijación de CO2 se necesitan 6 vueltas del
ciclo.
La clásica ecuación para la
síntesis de una hexosa es:
luz
6CO2 + 12H2O
C6H12O6 + 6O2 +
6H2O (4)
En ella se produce la acumulación de 686
kcal/mol de hexosa. Esta energía es provista por 12
NADPH y 18 ATP, que en total representan 877 kcal/mol. Por lo
tanto, la eficiencia del
proceso alcanza al 78%.
 Ciclo C 4:
Ciclo C 4:
Además del ciclo de Calvin (que se halla en la
mayoría de las plantas superiores) en cierto
número de especies de angiospermas se encuentra el ciclo
C4. Desde el punto de vista de la biología celular es de
gran interés
que ambos ciclos, el C4 y el C3, se hallan integrados en la
misma planta.
En las células mesófilas se asimila el
CO2 por la carboxilación del
fosfoenolpiruvato (PEP), lo que da origen a los ácidos
C4, como aspartato y malato. El CO2 que sale por
descarboxilación entra en el ciclo de Calvin y da origen
al 3-fosfoglicerato, mientras que los productos C3 pueden
volver a la célula mesófila y entrar en el ciclo
C4.
 RESPIRACION CELULAR:
RESPIRACION CELULAR:
Al igual que las células animales, las
vegetales efectúan las reacciones de la
respiración celular, principalmente en sus mitocondrias,
utilizando para ello sustratos como la glucosa y produciendo
CO2. En estas reacciones hay producción de
ATP, el cual sirve para impulsar los procesos
metabólicos de las células. Además en
determinadas condiciones, muchas plantas utilizan
oxígeno y producen CO2 a través de un
proceso diferente llamado fotorrespiración.
En días de sol intenso, cuando el clima es
caliente y seco, las células vegetales cierran sus
estomas (pequeños poros de la superficie foliar) para
evitar la pérdida de agua a través de las hojas.
Esta respuesta impide la deshidratación pero al mismo
tiempo el paso de CO2 al interior de la hoja. Al
llevar a cabo la fotosíntesis en estas condiciones, el
CO2 es expulsado y la concentración de
oxígeno aumenta. Cuando la concentración de
oxígeno en la hoja es mayor que la concentración
de CO2, el oxígeno se combina en vez del
CO2, con el sitio activo de la ribulosa difosfato
carboxilasa. Esta enzima es una oxigenasa y al mismo tiempo es
una carboxilasa; el CO2 y el carbono y el
oxígeno compiten entre sí por el sitio activo de
esta enzima. Cuando actúa como carboxilasa, cataliza la
combinación del CO2 con la ribulosa 1,5
difosfato y el oxígeno molecular, formando un compuesto
de tres carbonos, el 3-fosfoglicerato, y uno de dos carbonos,
el 2-fosfoglicolato. El fosfoglicolato se hidroliza, formando
el glicolato y fosfato inorgánico. Luego el glicolato
abandona el cloroplasto y más adelante se metaboliza en
un glioxisoma. El glicolato se convierte en glioxilato,
liberando peróxido de hidrógeno. Posteriormente
el glioxilato se metaboliza en las mitocondrias, formando
CO2.
La fotorrespiración consume hasta 50% del
CO2 fijado en el ciclo de Calvin; por tanto parece
ser un proceso de desgaste sin beneficio aparente para la
planta. Si pudiera disminuirse la fotorrespiración,
podría aumentarse el abastecimiento de alimento.
Nótese que las plantas C4 fijan CO2 en
condiciones que provocarían la fotorrespiración
en las plantas C3.
2.2.7.1.- ÁCIDOS
NUCLEICOS
2.2.7.1.1.- Estructura molecular.-
Deseamos proponer una estructura para la sal del
ácido desoxirribonucleico (ADN). Esta estructura presenta
características nuevas que son de considerable
interés biológico.
Pauling y Corey ya han propuesto una estructura para el
ácido nucleico. Con gentileza nos brindaron su manuscrito
aun antes de publicarlo. Su modelo consta
de tres cadenas entrelazadas, cuyos fosfatos se encuentran cerca
del eje de fibra, mientras que, las bases dan al exterior. En
nuestra opinión, esa estructura es insatisfactoria por dos
razones: (1) Creemos que el material que produce los diagramas de
rayos X es la
sal, no el ácido libre. Sin los átomos de
hidrógeno acídicos no esta claro que fuerzas
podrían mantener unida la estructura, de manera especial
dado que los fosfatos cargados negativamente que hay cerca del
eje se repelerían unos a otros; (2) Algunas de las
distancias de Van der Waals parecen demasiado
pequeñas.
Fraser (en prensa) ha
propuesto también otra estructura de tres cadenas. En su
modelo, los fosfatos están por fuera y las bases en el
interior , unidas por enlaces de hidrógeno. Es una
estructura algo deficientemente definida, cual se describe, y por
lo mismo no la comentaremos.
Deseamos proponer una estructura radicalmente diferente
para la sal del ácido desoxirribonucleico. En una
estructura de dos cadenas helicoidales enrollada en torno al mismo
eje. Hemos hecho los supuestos químicos usuales, a saber,
que cada cadena consta de grupos diésteres de fosfasto
unidos a residuos de b-D-Desoxirribofuranosa con enlaces de 3',
5'. Ambas cadenas (que no las bases) se encuentran relacionadas
por una diada perpendicular al eje de fibra. Las dos cadenas
siguen un giro hacia la derecha, pero por la diada las secuencias
de loa átomos de ambas cadenas corren en dirección
contraria. Cada cadena se parece algo al modelo N° 1 de
Furberg, o sea, las bases están en el interior de la
hélice y los fosfatos por fuera . La configuración
del azúcar y de los átomos cercanos a él se
acerca a la "configuración estándar" de Furberg,
con el azúcar más o menos perpendicular a la base
que lleva adherida. En cada cadena hay un residuo cada 3-4 A en
dirección -z. Hemos supuesto un ángulo de 36°
entre residuos ayacentes en la misma cadena, de manera que la
estructura se repite en cada cadena después de 10
residuos, o sea, después de 34 A. La distancia de un
átomo
de fósforo al eje de fibra es de 10 A. Como los fosfatos
están en el exterior, los cationes les llegan
fácilmente.
Es una estructura abierta y su contenido de agua es
más bien alto. Si el de agua fuera inferior cabría
esperar que las bases se inclinaran, de donde la estructura
resultaría más compacta.
La estructura tiene de característica novel el
modo como se mantienen las dos cadenas por las bases
purínicas y pirimidínicas . Los planos de las bases
son perpendiculares al eje de fibra. Van en pares, de forma que
cada base de una cadena se enlaza por medio de hidrógeno
con otra base de otra cadena, una al lado de otra con
idénticas coordenadas -z. Para que pueda ocurrir el
enlace, un elemento del par ha de ser de una purina y el otro de
una pirimidina. Los enlaces de hidrógeno son como sigue:
posición purínica 1 a posición
pirimidínica 1; posición purínica 6 a
posición pirimidínica 6.
Si se supone que las bases sólo ocurren en la
estructura en las formas tautoméricas más
plausibles (o sea, con las configuraciones keto y no con la enol)
resulta que sólo se pueden enlazar pares
específicos de bases . Dichos pares: adenina (purina) con
timina (pirimidina), y guan (purina) citosina
(pirimidina).
En otras palabras, si una adenina forma un miembro del
par, en ambas cadenas, entonces de tal supuesto el otro miembro
ha de ser timina, y lo mismo vale para la guanina y la citosina.
La secuencia de bases de una cadena no parece tener
restricción de ningún tipo. Sin embargo, si solo se
pueden formar pares específicos de bases, se sigue que
sabiendo la secuencia de bases de una cadena se puede determinar
automáticamente la secuencia en la otra cadena.
Se ha visto experimentalmente que la proporción
en la cantidad de adenina frente a la timina, y la
proporción de guanina a citosina, son siempre muy
próximas a la unidad en el ácido
desoxirribonucleico.
Quizá sea imposible construir esta estructura con
un azúcar de ribosa en vez de la desoxirribosa, puesto que
el átomo de oxigeno extra
acercaría demasiado la distancia de Van der
Waals.
Los datos de rayos X
antes publicados sobre el ácido desoxirribonucleico para
comprobar con rigor nuestra estructura. Hasta donde podemos
afirmar es compatible en general con los datos experimentales,
pero se ha de considerar como no probada hasta compulsarla con
resultados más exactos. En las siguientes comunicaciones
se presentan algunos de éstos. No nos habíamos
percatado de los detalles de los resultados allí
presentados, al idear la estructura , que se basa principalmente
en datos experimentales publicados ( aunque no del todo) y en
argumentos estereoquímicos.
No se nos ha escapado que el apareamiento
específico que hemos postulado sugiere de inmediato la
existencia de un posible mecanismo de copiado de material
genético.
Los detalles completos de la estructura , incluidas las
condiciones supuestas para construirla, junto con un par de
coordenadas de los átomos, se publicarán en otra
parte.
Estamos muy en deuda con el Dr. Jerry Donohue, por su
constante consejo y crítica, sobre todo respecto de las
distancias interatómicas . Nos ha estimulado asimismo el
haber tenido conocimiento,
de una manera general , de los resultados inéditos de
experimentos y
de ideas del Dr. M.H.F. Wilkins, de la Dra. R. E. Flanklin y sus
colaboradores en el King's College, Londres. Uno de nosotros (J.
D.W.) dispone de la ayuda de una beca de la Fundación
Nacional pro Parálisis Infantil.
2.2.7.2.-Síntesis de
proteínas
La traducción del ARNm
2.2.7.2.1.-INTRODUCCION
El ARN mensajero es el que lleva la información
para la síntesis de proteínas, es decir, determina
el orden en que se unirán los
aminoácidos
La síntesis de proteínas o
traducción tiene lugar en los ribosomas del
citoplasma celular. Los aminoácidos son transportados por
el ARN de transferencia (ARNt) , específico para cada uno
de ellos, y son llevados hasta el ARN mensajero (ARNm),
dónde se aparean el codón de éste y
el anticodón del ARN de transferencia, por
complementariedad de bases, y de ésta forma se
sitúan en la posición que les
corresponde.
Una vez finalizada la síntesis de una
proteína, el ARN mensajero queda libre y puede ser
leído de nuevo. De hecho, es muy frecuente que
antes de
que finalice una proteínaya está comenzando
otra, con lo cual, una misma molécula de ARN mensajero,
está siendo utilizada por varios ribosomas
simultáneamente.
- Los ARNt desempeñan un papel central en la
síntesis de las proteínas
La síntesis proteica tiene lugar en el ribosoma,
que se arma en el citosol a partir de dos subunidades
riborrucleoproteicas provenientes del nucléolo. En el
ribosoma el ARN mensajero (ARNm) se traduce en una
proteína, para lo cual se requiere también la
intervención de los ARN de transferencia (ARNt). El trabajo de
los ARNt consiste en tomar del citosol a los aminoácidos y
conducirlos al ribosoma en el orden marcado por los
nucleótidos
del ARNm, que son los moldes del
sistema
La síntesis de las proteínas comienza con
la unión entre sí de dos aminoácidos y
continúa por el agregado de nuevos aminoácidos -de
a uno por vez- en uno extremos de la cadena.
Como se sabe la clave de la traducción reside en
el
código genético, compuesto por
combinaciones de tres nucleótidos consecutivos -o
tripletes- en el ARNm. Los distintos tripletes se relacionan
específicamente con tipos de aminoácidos usados en
la síntesis de las proteínas.
Cada triplete constituye un codón: existen en
total 64 codones, 61 de los cuales sirven para cifrar
aminoácidos y 3 para marcar el cese de la
traducción. Tal cantidad deriva de una relación
matemática
simple: los cuatro nucleótidos (A, U, C y G)se combinan de
a tres, por lo que pueden generarse 64
(43).
Dado que existen más codones, (61) que tipos de
aminoácidos (20), casi todos pueden ser reconocidos por
más de un codón, por lo que algunos tripletes a
como "sinónimos". Solamente el triptófano y la
metionina -dos de los aminoácidos menos frecuentes en las
proteínas - son codificados, cada uno, por un solo
codón.
Generalmente los codones que representan a un mismo
aminoácido se parecen entre sí y es frecuente que
difieran sólo en el tercer nucleótido. La baja
especificidad de este nucleótido ha llevado a decir que
existe
una "degeneración" en tercera base de la
mayoría de los codones. Resta agregar que el número
de codones en el ARNm determina la longitud de la
proteína.
- Existen 31 tipos diferentes de
ARNt
Las moléculas intermediarias entre los codones
del ARNm y los aminoácidos son los ARNt, los cuales tienen
un dominio que se
liga específicamente a uno de los 20 arninoácidos y
otro que lo hace, específicamente también, con el
codón apropiado. El segundo dominio consta de una
combinación de tres nucleótidos -llamada
anticodón - que es complementaria de la del
codón.
Cada tipo de ARNt lleva antepuesto el nombre del
aminoácido que transporta. por ejemplo, leucinil-ARNt para
el aminoacil-ARNt de la leucina, lisinil-ARNt para el de la
lisina, fenilalanil-ARNt para el de la fenilalanina,
metionil-ARNt para el de la metionina,
etcétera.
Por su lado. El ARNt unido al aminoácido
compatible con él se designa aminoacil-ARNtAA,
en el que "AA" correspnde a la sigla del aminoácido. Por
ejemplo, leucinil-ARNtLeu, lisinil-ARNtlys,
fenilalanil-ARNtPhe. metionil-ARNtMet,
etcétera.
Si bien teóricamente pueden existir 61 tipos de
ARNt diferentes, sólo hay 31. El déficit se
resuelve por la capacidad que tienen algunos ARNt de reconocer a
más de un codón. Lo logran porque sus anticodones
suelen poseer la primera base
"adaptable", es decir, que
puede unirse con una base no complementaria situada en la tercera
posición del codón (recuérdese la
"degeneración" de esta base).
Así, la G en la primera posición del
anticodón puede aparearse tanto con una C -es lo habitual
- como con una U del codón . Similarmente, la U en la
primera posición del anticodón puede hacerlo con
una A -es lo habitual - o con una G. Por otra parte, la inosina
(I) -una de las bases inusuales se encuentra en la primera
posición del anticodón en varios ARNt y es capaz de
aparearse con cualquier base (excepto con una G) localizada en la
tercera posición del codón.
- El codón de iniciación es el
triplete AUG
El primer codón que se traduce en los ARNm es
siempre el triplete AUG. cuya información codifica al
aminoácido
metionina . Por lo tanto, este
codón cumple dos funciones: señala el sitio de
comienzo de la traducción -caso en el cual recibe el
nombre de codón de iniciación -, y cuando se halla
en otras localizaciones en el ARNm codifica a las metioninas del
interior de las moléculas proteicas.
Al especificar el primer aminoácido de la
proteína, el codón AUG de iniciación
determina el encuadre de los sucesivos tripletes, lo que asegura
la síntesis correcta de la molécula. Tómese
como ejemplo la secuencia AUGGCCUGUAACGGU. Si el ARNm es
traducido a partir del codón AUG, los codones
siguientes serán GCC, UGU, AAC y GGU, que
codifican, respectivamente, a los aminoácidos alanina,
cisteina ,asparagina y glicina. En cambio, si se omitiera la A
del codón de iniciación, el encuadre de los
tripletes sería el siguiente: UGG, CCU, GUA y ACG, los
cuales se traducen en los aminoácidos triptófano,
prolina, valina y treonina, respectivamente.
Algo semejante ocurriría si también se
omitiera la U, pues resultaría un tercer tipo de encuadre:
GGC, CUG, UAA y CGC. En este caso, después de codificar
los dos primeros codones a los aminoácidos glicina y
leucina, la traducción se detendría, ya que UAA es
un codón de terminación.
- Los aminoácidos se ligan por medio de
uniones peptídicas
La unión de los aminoácidos entre
sí para construir una proteína se produce de modo
que el grupo carboxilo de un aminoácido se combina con el
grupo a amínoácido siguiente, con pérdida de
una molécula de agua H2O y recordemos que esa
combinación se llama unión
peptídica.
Cualquiera que sea su longitud, la proteína
mantiene el carácter
anfotérico de los aminoácidos aislados, ya que
contiene un grupo amino libre en uno de sus extremos y un grupo
carboxilo en el otro extremo. La proteína se sintetiza a
partir de extremo que lleva el grupo amino libre. Ello se
corresponde con la dirección 5´ 3´
usada para la traducción del ARNm, la misma con que el ADN
se transcribe.
Antes de describir los procesos que dan lugar a la
síntesis de las proteínas analizaremos cómo
arriban los ARNm al citoplasma, qué configuración
poseen los ARNt y cuál es la estructura de los
ribosomas.
- Los ARNm arribados al citoplasma se conectan con
ríbosomas
Los transcriptos primarios de los ARNm se hallan
combinados con diversas proteínas, con las que forman las
nueleoproteínas heterogéneas nucleares o RNPhn.. No
obstante, muchas de esas proteínas se desprenden de los
ARNm a medida que éstos abandonan el
núcleo.
Los ARNm salen hacia el citoplasma por los poros de la
envoltura nuclear. Ya en el citosol, cada ARNm se combina con
nuevas proteínas y con ribosomas, lo que lo habilita para
ejercer su función codificadora durante la síntesis
proteica. Entre las proteínas se encuentra la llamada CBP
(por cap
binding protein), que se combina con el cap en el
extremo 5´ del ARNm.
Algunos ARNm se localizan en sitios prefijados en el
citoplasma, de modo que las proteínas que codifican se
sintetizan y se concentran en esos sitios. Un ejemplo es el ARNm
de la actina, que se sitúa en la zona periférica de
las células epiteliales donde se deposita la mayor parte
de la actina .
El extremo 5' de los ARNm contiene una secuencia de
alrededor de 10 nucleótidos previa al codón de
iniciación -entre éste y el cap - que, como es
lógico, no se traduce . En algunos ARNm esta secuencia
participa en el control de 1a traducción y en otros regula
la estabilidad del ARNm, es decir, su supervivencia.
Otra secuencia especial del ARNm, de hasta miles de
nucleótidos, suele hallarse después del
codón de terminación. entre éste y la poli A
. Tiene por función controlar la supervivencia del
ARNm.
- Las moléculas de los ARNt adquieren una
forma característica
Hemos visto que los codones del ARNm no seleccionan a
los aminoácidos directamente y que la traducción de
los ARNM en proteínas depende de un conjunto de
moléculas intermediarias -los ARNt- que actúan como
adaptadores, ya que discriminan tanto a los codones del ARNm como
a los aminoácidos compatibles con ellos.
Así la función básica de los ARNt
es alinear a los aminoácidos siguiendo el orden de los
codones para poder cumplir
con sus funciones, los ARNt ,adquieren una forma
característica semejante a un
trébol de cuatro
hojas . Los cuatro brazo
s se generan por la
presencia en los ARNt de secuencias de 3 a 5 pares de
nuelcótidos complementarios, los cuales se aparean entre
sí como los nucleótidos de las dos cadenas del
ADN.
En la punta de uno de los brazos confluyen los extremos
5' y 3´ del ARNt. El extremo 3´ es más largo,
de modo que sobresale el trinucleótido CCA que fue
incorporado durante el procesamiento. Este brazo se llama
aceptador porque a él se liga el aminoácido, que se
une a la A del CCA.
Los tres brazos restantes poseen en sus extremos
secuencias de 7 a 8 nucleótidos no apareados, -con forma
de asas -, cuyas denominaciones derivan de los nucleótidos
que las caracterizan. Una de ellas contiene el triplete de
nueleótidos del anticodón, por lo que su
composición varía en cada tipo de ARNt. Otra, en
virtud de que contiene dihidrouridinas (D), se denomina asa D. La
tercera se conoce como asa T, por el trinucleótido
T C que la identifica. La letra T simboliza a la
ribotimidina y la a la seudouri dina.
Entre el asa T y el anticodón existe un asa
adicional, llamada variable porque su longitud difiere en los
distintos ARN de transferencia.
Un plegamiento ulterior en el ARNt hace que deje de
parecerse a un trébol de cuatro hojas y adquiera la forma
de la
letra L . El cambio se debe a que se establecen
apareamientos inusuales entre algunos nueleótidos, como la
combinación de un nucleótido con dos a la
vez.
Formada la L, las asas D y T pasan a la zona de
unión de sus dos ramas y el brazo aceptador y el triplete
de bases del anticodón se sitúan en las puntas de
la molécula.
Una aminoacil-ARNt sintetasa une
el aminoácido al ARNt
El aminoácido se liga a su correspondiente ARNt
por la acción de una enzima llamada aminoacil-ARNt
sintetasa, que cataliza la unión en dos pasos.
Durante el primero, el aminoácido se liga a un
AMP , con el cual forma un aminoacil AMP. Por ejemplo leucinil
–AMP , lisinil AMP, fenilalanil AMP, metionil-AMP, etc..
Dado que el AMP deriva de la hidrólisis de un ATP , se
libera pirofosfato (PP) y energía , que también
pasa al aminoacil- AMP
AA + ATP AA-AMP + PP
En el segundo paso esa energía es utilizada por
la aminoacil ARNt sintetasa para transferir el aminoácido
del aminoacil –AMP a la A del brazo aceptador del ARNt
compatible, con lo cual se forma una molécula esencial
para la síntesis proteica: el aminoacil-ARNtAA
que reconoce el codón complementario en el
ARNm.
AA-A + ARNt (
AMINOACIL
SINTETASA) AA-ARNtAA +
AMP
Debe señalarse que la energía del ATP
usada en la primera reacción queda depositada en la
unión química entre el aminoácido y la A del
trinucleótido CCA.
- Existen 20 amínoacil – ARNt
sintetasas diferentes
Existen 20 aminoacil-ARNt sintetasas diferentes, cada
una diseñada para reconocer a un aminoácido y al
ARNt compatible con él. Ambos reconocimientos permiten
que cada uno de los 31 tipos de ARNt
se ligue sólo a uno de los 20
aminoácidos usados en la síntesis proteica. Ello
es posible porque cada aminoacil ARNt sintetasa identifica al
ARNt por el anticodón, la parte más
específica del ARNt . No obstante, en los ARNt existen
otras señales que son reconocidas por la enzima,
generalmente tramos de nucleótidos cercanos al
anticodón.
Como es obvio, la existencia de 11 clases de ARNt hace
que algunos aminoácidos sean reconocidos por más
de un ARNt.
Uno de los ARNt redundantes es el llamado ARNt
iniciador o ARNt[i], pues transporta a la metionina destinada
exclusivamente al codón AUG de iniciación . Es
muy probable quecerca de ese codón existan
señales que diferencien al
metionil-ARNt[i]met –portador de la metionina
dirigida a él- de los metionil ARNtmet
comunes, portadores de las metioninas destinadas a los
restantes codones AUG del ARNm.
- Los ribosomas están compuestos por dos
subunidades
Los mecanismos para alinear a los aminoacil
ARNtAA de acuerdo con el orden de los codones del
ARNm son algo complicados. Requieren de los ribosomas cuya
primera tarea es localizar al codón AUG de
iniciación y acomodarlo correctamente para que el
encuadre de ese triplete y el de los siguientes sea el
adecuado.
Luego el ribosoma se desliza hacia el extremo
3´del ARNm y traduce a los sucesivos tripletes en
aminoácidos. Estos son traídos – de a uno
por vez – por los respectivos ARNt. Las reacciones que
ligan a los aminoácidos entre sí - es decir , las
uniones peptídicas - se producen dentro del ribosoma .
Finalmente, cuando el ribosoma arriba al codón de
terminación – en el extremo 3´del ARNm
– cesa la síntesis proteica y se libera la
proteína. Como podemos notar, los ribosomas constituyen
las "
fábricas de las proteínas"
Cada ribosoma está compuesto por dos
subunidades - una mayor y otra menor – identificadas con
las siglas 40S y 60S respectivamente (los números hacen
referencia a los coeficientes de sedimentación de las
subunidades, es decir a las velocidades con que sedimentan
cuando son ultracentrifugadas, la 60S migra más
rápido al fondo del tubo).
En la subunidad menor algunas proteínas forman
dos áreas - una al lado de la otra – denominadas
sitio P (por
peptidil) y sitio A (por
aminoacil).
Por otro lado en la subunidad mayor las
proteínas ribosómicas formarían un
túnel por el que saldría la cadena
polipeptídica a medida que se sintetiza
Las etapas de la síntesis de
proteínas
La síntesis de las proteínas se divide en
tres etapas, llamadas de
iniciación , de
alargamiento y de
terminación.
- El comienzo de la síntesis proteica
requiere de varios factores de
iniciación
La etapa de
iniciación es regulada por
proteínas citosólicas denominadas factores de
iníciación (IF), que provocan dos hechos separados
pero concurrentes , uno en el extremo 5´del ARNm y otro en
la subunidad menor del ribosoma
El primer proceso involucra al cap y a una secuencia de
nucleótidos aledaña, localizada entre el cap y el
codón de iniciación . Estas partes reconocidas por
el factor IF-4, que se liga a ellas sí al ARNm se
proteína CBP . La conexión del IF-4 con el ARNm
insume energía que es provista por un ATP.
En el segundo proceso, el
metioníl-ARNt[i]met se coloca en el sitio P de
la subunidad menor del ribosoma, reacción que requiere el
factor IF-2 y la energía de un GTP.
Logrados ambos acondicionamientos, otro factor de
iniciación, el IF-3, con la ayuda del IF-4 coloca el
extremo 5´ del ARNm sobre una de las caras de la unidad
menor del ribosoma, la que posee los sitios P y A.
De inmediato la subunidad menor se desliza por el ARNm y
detecta al codón de AUG de iniciación, que se
coloca
, en el sitio P . Como es lógico , el segundo
codón del ARNm queda colocado al lado, es decir en el
sitio A.
Entre tanto, el metioril-ARNt[i]met ,'
ubicado en el sitio P de la subunidad menor, se une al
codón AUG de iniciación mediante su
anticodón CAU (UAC ). El acoplamiento correcto
entre estos dos tripletes es imprescindible para asegurar el
encuadre normal de los siguientes codones del ARNm en los sitios
P y A del ribosoma.
La etapa de iniciación concluye cuando la
subunidad menor se combina con la subunidad mayor y se forma el
ribosoma. En él se encuentran los primeros dos codones del
ARNm: en el sitio P el codón AUG de iniciación
-unido al metionilARNt[i]met- y en el sitio A el
codón que le sigue.
La unión entre sí de las dos subunidades
ribosómicas se produce luego del desprendimiento del IF-2
y del IF-3, lo cual es mediado por el factor IF-5.
- El alargamiento de la cadena proteica es promovido
por factores de elongación
La
etapa de alargamiento comienza cuando al sitio
A del ribosoma se acerca otro aminoacil-ARNtAA,
compatible con el segundo codón del ARNm, con el cual se
une. La reacción es mediada por un factor de
elongación llamado EF-1 y consume energía, que es
aportada por un GTP.
Al quedar el aminoacil-ARNtAA cerca del
metionil-ARN[t]met. la metionina localizada en el
sitio P, al tiempo que se desacopla del. ARNt[i], se liga -
mediante una unión peptidica - al aminoácido
ubicado en el sitio A. Se forma así un dipeptidil-ARNt,
que continúa ubicado en el sitio A. Su permanencia en este
sitio es breve, en seguida veremos por qué.
La unión peptídica es catalizada por la
subunidad mayor del ribosoma. Debe agregarse que la
energía requerida para consumar esa unión proviene
de la ruptura de otra unión química , aquella que
liga al aminoácido con la adenina en el brazo aceptador
del ARNt. Como en el caso del metionil – ARNt
[i]met, la ruptura química tiene lugar siempre
en el sitio P.
Entre tanto, fuera del ribosoma, esperando para
ingresar, se encuentra el tercer codón del ARNm. Aborda el
ribosoma cuando el ARNm se corre tres nucleótidos en
dirección de su extremo 5´. Este proceso –
llamado traslocación – es mediado por el el factor
de elongación EF-2 y también consume energía
ahora aportada por un GTP.
Como vemos, desde el punto de vista energético la
síntesis proteica es bastante costosa, ya que por cada
aminoácido que se incorpora se consumen dos GTP y un ATP,
el último gastado durante 1a síntesis del
aminoacil-ARNtAA
El corrimiento del ARNm hace que el codón de
iniciación sea desalojado del sitio P sitio P -y, por
consiguiente, del ribosoma- el segundo codón se mude del
sitio A al sitio P y el tercer codón ingrese en el sitio A
vacante. Lógicamente el corrimiento de los codones
desplaza también a los ARNt , por lo que el ARNt[i] sale
del ribosoma -no tarda en desprenderse del codón de
iniciación – y el dipéptido pasa del sitio A
al sitio P.
Mientras tanto, un tercer aminoacil-ARNtAA
ingresa en le ribosoma , se acomoda en el sitio A y su
anticodón se une al tercer codón de ARNm, otra vez
por la intervención del EF-1. Debe señalarse que el
EF-1 actúa después que el EF-2 se retira del
ribosoma, y viceversa.
El paso siguiente comprende la formación de una
unión peptídica entre el dipéptido y el
aminoácido del tercer aminoacil –ARNt AA.
Esta unión peptídica, ahora entre e
dipéptido y el aminoácido del tercer
aminoacil-ARNtAA. Esta unión peptídica
genera un tripeptidil –AARNt, que permanece en el sitio P
hasta la próxima translocación del ARNm.
Los procesos citados se repiten de forma sucesiva
codón tras codón ; así , en el cuarto paso
se forma un tetrapeptidil ARNt y luego peptidil - ARNt cada vez
más largos , que se traslocan del sitio A al P conforme se
producen las uniones peptídicas. Se calcula que se agregan
a la cadena, en promedio, cinco aminoácidos por
segundo.
Debido a que con cada traslocación se corren tres
nucleótidos del ARNm , su extremo 5´se aleja
progresivamente del ribosoma y su extremo 3´se acerca a
él en igual medida. Cuando el ribosoma se ha alejado del
extremo 5´del ARNm unos 90 nucleótidos, en el
codón de iniciación se acomoda un nuevo ribosoma,
lo cual da inicio a la síntesis de otra cadena proteica.
Esto se repite varias veces .
- La síntesis proteica concluye cuando el
ribosoma alcanza el codón de
terminación
La etapa de terminación determina la
conclusión de la síntesis de la proteína
cuando el sitio A del ribosoma es abordado por el codón de
terminación del ARNm (UUA, UGA o UAG, indistintamente).
Ello deja al sitio A sin el esperado aminoacil-ARNtAA,
aunque pronto es ocupado por un factor de terminación
llamado eRF (eucaryotic releasing factor), que sabe reconocer a
los tres codones de terminación.
En síntesis la terminación de la cadena
polipeptídica está señalada por el ARNm
mediante un codón que no especifica la
incorporación de ningún aminoácido . Ese
codón de terminación puede ser UUA, UGA o UAG, y
sobre él no se une ningún ARNt. En cambio, es
reconocido por dos proteínas llamadas factores de
liberación (eRF). Cuando esto sucede, la proteína
terminada se libera del último ARNt, que también se
separa del ARNm. Por último también se disocian las
subunidades ribosómicas. Todos estos elementos pueden ser
reutilizados en una nueva síntesis.
2.2.8.- DIVISIÓN
CELULAR
2.2.8.1.- División celular en procariotas
|
Contenidos
Los
procariotas tienen una organización
mucho más simple que la de los eucariotas, los cuales
entre otras cosas, tienen muchos más cromosomas.
El cromosoma procariota es una sola molécula circular de
ADN contenida en una región definida del citoplasma,
denominada nucleoide, sin estar separado del mismo por una
membrana. Este cromosoma es el elemento obligatorio del
genoma, aunque es frecuente encontrar unidades
de replicación autónomas llamadas
plásmidos, que si se pierden, la bacteria
sigue siendo viable.
El método
usual de duplicación de las células procariotas se
denomina fisión binaria. La duplicación de la
célula va precedida por la
replicación
del cromosoma bacteriano. Primero se replica y luego pega
cada copia a una parte diferente de la membrana celular. Cuando
las células que se originan comienzan a separarse,
también se separa el cromosoma original del
replicado.
Luego de la separación (
citocinesis),
queda como resultado dos células de idéntica
composición genética (excepto por la posibilidad de
una mutación espontánea)
Una consecuencia de este método asexual de
reproducción es que todos los organismos de una colonia
son genéticamente iguales. Cuando se trata una enfermedad
originada en una infección bacteriana, una droga que mata
a una bacteria matará a todos los miembros de ese clon
(colonia).
2.2.8.2.- División celular en eucariotas
|
Contenidos
En razón de su número de cromosomas,
organelas y complejidad la división de la célula
eucariota es más complicada, aunque ocurran los mismos
procesos de replicación, segregación y
citocinesis.
2.2.8.3.- Mitosis |
Contenidos
La
mitosis es el proceso de formación de dos
células idénticas (generalmente) por
replicación y división de los cromosomas de la
original que da como resultado una "copia" de la
misma.
Las células eucariotas poseen un mayor
número de cromosomas que por otra parte son mucho
más grandes que los de los procariotas.
Los estructura de los cromosomas replicados y
condensados tiene varios aspectos de interés.
El cinetocoro es el punto donde "anclan" los
microtúbulos del huso. Los cromosomas replicados consisten
en dos moléculas de ADN (junto con sus proteínas
asociadas: las
histonas) que se conocen con el nombre de
cromátidas. El área donde ambas cromátidas
se encuentran en contacto se conoce como centrómero, el
cinetocoro se encuentra en la parte externa del
centrómero. Se debe hacer hincapié en que los
cromosomas son
cromatina (ADN más histonas) y
señalar la particularidad que en los extremos del
cromosoma (que toman el nombre de telómero) se encuentran
secuencias repetidas de ADN.
Dependiendo de la posición del centrómero
los cromosomas se clasifican en:
- telocéntricos, con el centrómero en un
extremo
- acrocéntricos, uno de sus brazos es muy
corto
- submetacéntricos, brazos de diferente
longitiud
- metacéntricos, brazos de igual
longitud
2.2.8.4.- Empaquetamiento del ADN |
Contenidos
Las proteínas asociadas al ADN se conocen
colectivamente con el nombre de histonas. Son polipéptidos
relativamente cortos cargados positivamente (básicos) y
por lo tanto son atraídos por las cargas negativas del ADN
(ácido) Las histonas son sintetizadas en cantidad
durante la fase S ( S por síntesis) del ciclo celular. Una
de las funciones de esas proteínas está relacionada
con el empaquetamiento del ADN en la forma del cromosoma: los 2
metros de ADN de la célula humana son empaquetados en 46
cromosomas de un largo combinado de aproximadamente 200 nm. La
célula tiene unas 90 millones de moléculas de
histonas siendo la mayoría perteneciente a un tipo
conocido como H1. Se conocen cinco tipos de las siguientes
histonas (H1, H2A, H2B, H3, y H4 , 8 moléculas en total);
con la excepción de la H1 la mayor parte de las histonas
de los eucariotas son muy similares.
El nucleosoma es la unidad fundamental de
"empaquetamiento" del ADN eucariótico. El "carretel"
("
core") del mismo consiste en dos moléculas de
H2A, H2B, H3, y H4; alrededor de las cuales el ADN se enrolla dos
veces . La histona 1 esta fuera del "carretel". Este nivel de
empaquetamiento ("
packing") se conoce como "cuentas de un
collar" . El siguiente nivel se conoce como la fibra de 30 nm,
cuyos detalles de organización no se conocen
completamente. Las fibras se condensa
a posteriori en
dominios en bucle de 300 nm . Los dominios son parte de las
secciones condensadas ( 700 nm) de los cromosomas (el cromosoma
tiene un ancho de unos 1.400 nm en la metafase) .
Durante la mitosis los cromosomas replicados se
posicionan cerca de la mitad de la célula y luego
se
segregan en manera tal que cada
célula resultante recibe una copia de cada cromosoma
original (si se comienza con 46 cromosomas en la célula
original se termina con 46 cromosomas en las 2 células
resultantes). Para realizar esto las células utilizan
microtúbulos (que en este caso en conjunto forman el huso
mitótico) que "tiran" de los cromosomas para llevarlos a
cada futura célula. Las células animales (excepto
un grupo de gusanos conocidos con el nombre de nematodos)
poseen
centríolos. Las plantas y la mayor
parte de los otros eucariotas no poseen centríolos y los
procariotas, por supuesto, carecen de huso y centríolos;
en procariotas la membrana celular suple esta función al
arrastrar los cromosomas pegados a ella durante la citocinesis de
la fisión binaria. Las células que contienen
centríolos también poseen una "corona" de
pequeños microtúbulos, el aster, que se extienden
desde los centríolos a la membrana nuclear.
Las fases de la mitosis son en realidad difíciles
de separar. Se debe tener en cuenta que el proceso no es el
estático que se describe en el texto, sino
dinámico como el que se puede seguir en
esta animación.
2.2.8.5.- Profase |
Contenidos
La profase es el primer estadio de la
mitosis. La cromatina se condensa (recordar que el ADN de la
cromatina se replica en la interfase), por lo que en este punto
existen dos cromátidas unidas. La membrana nuclear se
disuelve, los centríolos (si se encuentran presentes) se
dividen y los pares migran a los polos, se forma el huso
mitótico. Los centrómeros (o constricciones
primarias) se vuelven claramente visibles, debido a que se le han
asociados placas proteicas a ambos lados: el
cinetocoro. En el citoplasma el
retículo endoplasmático y el complejo de Golgi se
fragmentan en vesículas, se desorganiza el citoesqueleto
por lo que la célula pierde su forma original y se hace
esférica.
2.2.8.6.- Metafase |
Contenidos
La metafase sigue a la profase. Los cromosomas (que a
este punto consisten en dos cromátidas mantenidas juntas
por el centrómero) alcanzan su máxima
condensación y migran al ecuador de la
célula donde las fibras del huso se "pegan" a las fibras
del cinetocoro.
2.2.8.7.- Anafase |
Contenidos
La anafase comienza con la separación de los
centrómeros y el arrastre de las cromátidas (los
llamamos cromosomas luego de la separación de los
centrómeros) a los polos opuestos.
2.2.8.8.- Telofase |
Contenidos
En la telofase los cromosomas llegan a los polos de sus
respectivos husos, la membrana nuclear se reconstituye, los
cromosomas se desenrollan y pasan a formar la cromatina y el
nucleolo, que desapareció en la profase se vuelve a
constituir. Donde antes había una célula ahora
existen dos pequeñas con exactamente la misma
información genética y número
cromosómico. Estas células pueden luego
diferenciarse en diferentes formas durante el
desarrollo.
2.2.8.9.- Citocinesis |
Contenidos
La citocinesis es el proceso de separación de las
células formadas. En tanto la mitosis es la
división del núcleo en la citocinesis ocurre la
división y la relocalización de los
plástidos, Golgi y citoplasma en cada nueva célula.
Se reestablece el citoesqueleto.
Difiere en las células animales y vegetales. En
las primeras, la membrana comienza a constreñirse
alrededor de la circunferencia de la célula,
formándose un anillo contráctil de miosina y
actina.
En las células vegetales una serie de vesículas
producidas por los dictiosomas divide al citoplasma en la
línea media formando una placa celular que crece en forma
centrífuga y se fusiona a la membrana de la célula
madre dividiendo la célula en dos.
2.3.-
MICROBIOLOGIA.-
2.3.1.-. BACTERIAS/Morfología y
estructura.
Las bacterias son microorganismos procariotas de
organización muy sencilla. La célula bacteriana
consta:
- citoplasma. Presenta un aspecto viscoso, y
en su zona central aparece un nucleoide que contiene la mayor
parte del ADN bacteriano, y en algunas bacterias aparecen
fragmentos circulares de ADN con información
genética , dispersos por el citoplasma: son los
plásmidos.
La membrana plasmática presenta invaginaciones,
que son los
mesosomas, donde se encuentran enzimas que
intervienen en la
síntesis
de ATP, y los pigmentos fotosintéticos
en el caso de bacterias fotosintéticas.
En el citoplasma se encuentran
inclusiones de
diversa naturaleza
química.
Muchas bacterias pueden presentar
flagelos
generalmente rígidos, implantados en la membrana
mediante un
corpúsculo basal . Pueden poseer
también,
fimbrias o pili muy numerosos y cortos,
que pueden servir como pelos sexuales para el paso de ADN de
una célula a otra
Poseen ARN y
ribosomas característicos,
para la
síntesis
de proteinas.
- pared celular es rígida y con
moléculas exclusivas de bacterias.
2.3.2.-Nutrición
El éxito evolutivo de las bacterias se
debe en parte a su versatilidad metabólica. Todos los
mecanismos posibles de obtención de materia y
energía podemos encontrarlos en las
bacterias.
Según la fuente de carbono que utilizan, los
seres vivos se dividen en autótrofos, cuya principal
fuente de carbono es el CO2 , y heterótrofos cuando su
fuente de carbono es materia orgánica.
Por otra parte según la fuente de
energía, los seres vivos pueden ser fototrofos, cuya
principal fuente de energía es la luz, y los
organismos quimiotrofos, cuya fuente de energía es un
compuesto químico que se oxida.
Atendiendo a las anteriores categorías, entre
las bacterias podemos encontrar las siguientes formas, como
puede apreciarse en el esquema:
1.-Las bacterias
quimioheterótrofas,
utilizan un compuesto químico como fuente de carbono
, y a su vez, este mismo compuesto es la fuente de
energía.
2.- La mayor parte de las bacterias cultivadas en
laboratorios y las bacterias patógenas son de este
grupo.
- Las bacterias quimioautótrofas,
utilizan compuestos inorgánicos reducidos como fuente
de energía y el CO2 como fuente de carbono. Como por
ejemplo, Nitrobacter, Thiobacillus.
- Las bacterias fotoautótrofas,
utilizan la luz como fuente de energía y el CO2 como
fuente de carbono. Bacterias purpureas.
- Las bacterias fotoheterótrofas,
utilizan la luz como fuente de energía y
biomoléculas como fuente de carbono. Ejemplos como
Rodospirillum y Cloroflexus.
2.3.3.- Reproducción
Generalmente las bacterias se reproducen por
bipartición.
Tras la duplicación del ADN, que esta dirigida por la
ADN-polimerasa que se encuentra en los mesosomas, la pared
bacteriana crece hasta formar un tabique transversal separador
de las dos nuevas bacterias.
Pero además de este tipo de reproducción
asexual, las bacterias poseen unos mecanismos de
reproducción sexual o parasexual, mediante los cuales se
intercambian fragmentos de ADN .
Puede realizarse por :
- TRANSFORMACION: Consiste en el intercambio
genético producido cuando una bacteria es capaz de
captar fragmentos de ADN, de otra bacteria que se encuentran
dispersos en el medio donde vive.
- CONJUGACIÓN: En este proceso, una bacteria
donadora F+ transmite a través de un puente o pili, un
fragmento de ADN, a otra bacteria receptora F-. La bacteria
que se llama F+ posee un plásmido, además del
cromosoma bacteriano.
Puedes verlo en el esquema siguiente y su
correspondiente animación.
- TRANSDUCCIÓN: En este caso la transferencia
de ADN de una bacteria a otra , se realiza a través de
un virus bacteriófago, que se comporta como un
vector intermediario entre las dos
bacterias.
2.3.2.- HONGOS .
Se puede decir que hay hongos por todas partes y en
todos los medios.
En las aguas, en el suelo, en el aire (esporas),
parásitos en plantas, empleados en la industria
alimenticia y farmacéutica, en las raíces (y
líquenes simbióticos), en los prados y bosques
(setas).
Son hongos los mohos que surgen al pudrirse cualquier
materia orgánica, así como los causantes de
plagas en la agricultura
(es el caso del Cornezuelo del Centeno), y de enfermedades de la piel
como son las micosis cutáneas. También lo son la
levadura que se usa para la fabricación del pan y la
cerveza, los
que dan aromas a los quesos y los que se utilizan para la
obtención de la penicilina, como "Penicilium nonatum".
Así, el hombre
continuamente está conviviendo, padeciendo y disfrutando
de los hongos y sus consecuencias.
Existen hongos de todos los tamaños, desde los
microscópicos hasta las grandes setas, y de todos los
colores y
formas. Pero la característica común a todos
ellos es la ausencia de clorofila u otros pigmentos
fotosintéticos. Por ello, los hongos tienen necesidad de
encontrar las sustancias nutritivas ya elaboradas. Son, por
ello, heterótrofos y pueden ser saprofítos (que
se alimentan de sustancias en descomposición),
parásitos o simbióticos (en combinación
con otras plantas).
Los hongos están compuestos por filamentos
(hifas) que son hileras de células, que forman una red o
micelio.
Se reproducen por esporas que se forman en los
aparatos esporíferos, que es la parte más visible
del hongo.
Existen más de 50.000 especies.
Tradicionalmente se ha incluido a los hongos dentro
del reino vegetal, considerándolos, como plantas sin
clorofila, llamandoles también Micófitos y
Eumicetes. Sin embargo los hongos no son ni plantas ni
animales, sino otro reino distinto.
Las diferencias más visibles son las
siguientes:
- Las plantas se alimentan mayormente por medio de la
fotosíntesis.
- Los hongos se alimentan por
absorción.
- Los animales se alimentan por
ingestión.
Por lo tanto los micólogos (los que estudian
los hongos), rechazan su inclusión dentro de los
vegetales.
Los hongos se desarrollan prefentemente en lugares
húmedos y oscuros ya que no necesitan de la luz para
sobrevivir. Son incapaces de producirse por si mismo los
compuestos
orgánicos que necesitan para sobrevivir, por eso
viven como parasitos de otros seres vivos, o en simbiosis con
las plantas formando las llamadas micorrizas con las
raices.
Los hongos podemos clasificarlos en grandes
grupos:
-
Ascomicetes: Hongos cuyas esporas se
producen en saquitos (ascas).
-
Basidiomicetes: Sus esporas se producen en
los basidios y comprende desde los tizones a las
setas.
-
Ficomicetes: Son hongos parecidos a las
algas, casi todos acuáticos, y mohos
negros.
-
Mixomicetes: Son organismos mitad hongo,
mitad animal, que suelen clasificarse aparte.
De estos tres grupos a los Basidiomicetes y a los
Ascomicetes se les denomina "Hongos superiores", y sobre ellos
profundizare por ser las especies más
conocidas.
2.3.2.1.-Partes de un Hongo
En el hongo hay que diferenciar dos partes
fundamentales: el cuerpo vegetativo y el cuerpo
reproductor.
El cuerpo vegetativo, que se encuentra bajo tierra,
está formado por unos filamentos llamados hifas que
pueden ser unicelulares (con una sucesión de nucleos), y
pluricelulares.
El conjunto de todas las hifas es el micelio. El es el
que se encarga de absorver las subtancias minerales del
suelo para alimento del hongo.
El micelio en realidad es el hongo, ya que
la
seta (a la que vulgarmente se llama hongo), es
su aparato
reproductor.
2.3.2.2.- Reproducción de los
hongos
Los hongos se reproducen por esporas. Los hongos
superiores poseen unas células madre localizadas en el
himenio, son las encargadas de producir las esporas. En el caso
de los
Basidiomicetes
a estas células madre se les denomina Basidios,
mientras que las células madre de los
Ascomicetes
son los Ascos.
Las esporas de los basidios y los ascos son lanzadas
al exterior para la propagación de la especie. Si la
espora se deposita en un lugar cuyas condiciones sean
favorables dará origen al micelio. Este crecerá
bajo tierra o entre la hojarasca, se ramificará y se
entremezclará con los micelios de otras esporas. En el
terreno donde la humeda y las condiciones del medio sean
más adecuadas crecerá una seta que portará
en su himenio los ascos o basidios que expulsarán al
exterior las esporas, dando lugar de nuevo al ciclo reproductor
del hongo.
2.3.2.4.- Las Setas Aragonesas
Se van ha indicar aquí algunas setas que se
pueden encontrar en Aragón, y antes de empezar una
advertencia: hay setas venenosas e incluso mortales.
No hay ninguna regla general para diferenciar una seta
venenosa de otra que no lo es. La única solución
es identificar cada seta antes de comerla y no tocar ni probar
ninguna que no se este seguro de cual
es.
Las setas (los hongos) tienen un papel fundamental en
la naturaleza ya que descomponen y hacen accesible para las
plantas el material que descomponen, por lo tanto hay que
tratar de no perjudicarlas innecesariamente.
Las setas se deben de cortar (no arrancar), por el pie
sin dañar el micelio que queda en el suelo.
Algunas de las setas que se pueden encontrar en
Aragón son las siguientes:
-
Falsa Oronja (Amanitas
muscaria).
- Oronja Mortal (Amanitas phalloides).
- Seta de San Jorge (Tricholoma georgii).
- Colmenilla (Morchella esculenta).
- Trufa Negra (Tuber nigrum).
- Calabaza (Boletus edulis)
- Champiñon Silvestre (Agaricus
campestris).
- Rovellón (Lactarius deliciosus).
- Setas de Pino (Tricholoma terreum).
- Clavaria (Ramaria aurea).
- Seta de Cardo (Pleurotus eryngii).
- Seta de Chopo (Agrocybe aegerita).
2.3.3.- PROTOZOOS. Fauna
Los protozoos son los animales más sencillos ya
que están formados por una sola célula. Por
tanto, son organismos unicelulares. Mediante su única
célula realizan todas las funciones vitales.
Los protozoos son abundantísimos y se
encuentran en todos los lugares de la tierra, en especial, en
los sitios húmedos. Son, frecuentemente,
parásitos sobre animales, plantas y sobre el hombre, y
pueden producir enfermedades.
Existen unas 50.000 especies de protozoos y pueden
vivir aislados o formando colonias.
1.-Respiración
La respiración la realizan a través de
la membrana celular y por las partículas de agua
absorbidas con el alimento.
2.- Vacuola pulsátil
La expulsión del gas
carbónico la hacen por las vesículas o vacuolas
pulsátiles.
Cuando la vacuola pulsátil está llena de
agua, se abre y lo libera al exterior.
3.-Locomoción
Los protozoos se mueven de diversas formas. Los
ciliados, somo el Paramecio, lo realizan mediante el movimiento
rítmico y rápido de los cilios.
Otros protozoos se desplazan mediante el rápido
movimiento del flagelo o los seudópodos.
4.- Alimentación
La alimentación suele
realizarse mediante la captura del alimento que penetra en el
citoplasma a través de una abertura de la
membrana.
En el citoplasma se forman vacuolas nutritivas y los
residuos son expulsados por las vacuolas fecales.
El paramecio succiona el alimento produciendo un
torbellino con los cilios.
Las amebas atrapan el alimento rodeándolo con
los seudópodos que forman.
5.- Reproducción
Los protozoos pueden reproducirse por
bipartición (división en dos), por
gemación (crecimiento de una yema o célula hija)
y por esporulación (fragmentación de la
célula madre en esporas). Cuando sucede este
último caso, pueden permanecer mucho tiempo enquistados
en una cápsula.
6.- Distintos tipo de Protozoos
6.1.- Flagelados
Los protozoos flagelados o mastigóforos
están provistos de uno o varios flagelos que les
permiten moverse.
Son unicelulares, se reproducen por división
longitudinal (a lo largo); viven libremente y muchos son
parásitos que producen enfermedades, algunas muy graves,
especialmente las tricomoniasis, la enfermedad del
sueño, la enfermedad de Chagas, la leptomoniasis,
etc.
En la clase de los flagelados se incluyen los
fitoflagelados
o dinoilagelados, que se tratan en el Reino
Vegetal (Algas unicelulares).
6.2.-Tripanosoma gambiensis
El tripanosoma es el causante de la enfermedad del
sueño, y es transmitida por la mosca
tsé-tsé. La mosca, al picar a un hombre
infectado, chupa algunos tripanosomas que se multiplican en su
intestino. Luego, cuando pica de nuevo, inyecta los protozoos
que se reproducen e invaden el cerebro.
6.3.-Trichomonas
Causante de varias enfermedades, en la boca,
intestino, vagina. etc
6.4.-Ciliados
Es la Clase más numerosa de los protozoos.
Poseen cilios en la membrana que usan para desplazarse.
Otros tienen cirros en forma de patas.
Muchos son libres, nadadores y otros viven sujetos por
pedúnculos que pueden enrollarse como un muelle.
Viven en aguas dulces o marinas.
Algunos son parásitos en peces
mamíferos e incluso en el hombre.
En periodos secos pueden enquistarse.
Pueden citarse entre otros al Paramecium, Vorticella Stentor,
Stilonichia, Balantidium, Mesódinium, etc.
6.5.- Suctores o Acinetos
Sólo presentan cilios en su estadio juvenil. De
adultos se fijan al sustrato mediante un pedúnculo y
poseen unos tentáculos huecos con ventosas, con las
cuales aferran a sus víctimas y succionan sus
jugos.
6.6.- Rizópodos o sarcodinos
Estos protozoos poseen seudópodos o
prolongaciones a modo de pies, para moverse y atrapar el
alimento.
Unos, como los Ameboides no tienen membrana rígida;
otros, los foraminíferos, radiolarios y heliozoos poseen
un esqueleto silíceo o calcáreo y sus
seudópodos son radiantes de infinidad de formas y
dibujos.
Existen en inmensas cantidades en mares y ríos, formando
parte importante del plancton.
6.7.-Esporozoos
Son todos parásitos que carecen de
órganos locomotores y digestivos.
Se reproducen por esporas resistentes, y también
sexualmente mediante la producción de un zigoto.
Este tipo de reproducción cíclica origina algunas
enfermedades graves, como las fiebres terciarias o paludismo.
ocasionada por el Plasmodium.
2.3.4.- ALGAS. Flora Silvestre.
Las Algas son los vegetales pluricelulares más
sencillos, ya que su estructura está formada por el
talo, que es una agrupación de células con cierta
diferenciación, similares a hojas, raíces o
tallos.
No poseen por lo tanto, tejidos, vasos conductores,
hojas ni raíces, pero ciertas partes de la planta asumen
funciones específicas.
Poseen plastos ricos en clorofila y otros
pigmentos.
La reproducción se realiza en fases alternas,
sexual y asexualmente.
Las algas pueden ser algas rojas o Rodofíceas;
algas pardas o Feofíceas y algas verdes o
Clorofíceas.
- El Conceptáculo masculino o anteridio produce:
Anterozoides o gametos masculinos
- El Conceptáculo femenino u Oogonio produce:
Oosferas o gametos femeninos
- El resultado es Huevos o Zigotos
- Mas adelante se desarrolla un
Embrión
- Al final aparece un Plántula
2.3.4.1.- CLASIFICACION DE LAS ALGAS
De menor a mayor complejidad podemos hacer la
siguiente clasificación:
Otras Informaciones de interes
2.3.5.- VIRUS ANIMALES Y
VEGETALES.-
Un virus es un agente genético que posee un
ácido nucléico que puede ser
ADN o
ARN, rodeado de una envuelta de
proteína.
Los virus contienen toda la información necesaria
para su ciclo reproductor; pero necesitan para conseguirlo a
otras células vivas de las que utilizan orgánulos y
moléculas.
Los virus pueden actuar de dos formas distintas:
- Reproduciéndose en el interior de la
célula infectada, utilizando todo el material y la
maquinaria de la célula hospedante.
- Uniéndose al material genético de la
célula en la que se aloja, produciendo cambios
genéticos en ella.
Por eso se pueden considerar los virus como agentes
infecciosos productores de enfermedades o como agentes
genéticos que alteran el material el material hereditario
de la célula huésped.
2.3.5.1.-REPRODUCCIÓN DE LOS VIRUS
La única función que poseen los virus y
que comparten con el resto de los seres vivos es la de
reproducirse o generar copias de sí mismos, necesitando
utilizar la materia, la energía y la maquinaria de la
célula huésped, por lo que se les denomina
parásitos obligados. No poseen metabolismo ni
organización celular, por lo que se les situa en el
límite entre lo vivo y lo inerte.
Los virus una vez infectan a una célula,pueden
desarrollar dos tipos de comportamiento, bien como agentes infecciosos
produciendo la lisis o muerte de la
célula o bien como virus atenuados, que añaden
material genético a la célula hospedante y por lo
tanto resultan agentes de la variabilidad
genética.
Ambos casos han sido estudiados con detalle en los virus
bacteriófagos, y aquí puedes ver en unos dibujos
esquemáticos:
En los dos casos de infección el proceso empieza
de esta forma:
- Fase de fijación : Los virus se unen
por la placa basal a la cubierta de la pared
bacteriana.
- Fase de contracción: La cola se contrae
y el ácido nucléico del virus se empieza a
inyectar.
- Fase de penetración : El ácido
nucléico del virus penetra en el citoplasma de la
bacteria, y a partir de este momento puede seguir dos ciclos
diferentes:
- En el ciclo lítico el ADN bacteriano fabrica
las proteínas víricas y copias de ácidos
nucléicos víricos. Cuando hay suficiente
cantidad de estas moléculas, se produce el ensamblaje de
la proteína y el A.N. vírico y se liberan al
medio, produciendo la muerte de
la célula.
- En el ciclo lisogénico se produce cuando el
genoma del virus queda integrado en el genoma de la bacteria,
no expresa sus genes y se replica junto al de la bacteria.
El virus queda en forma de profago.
3.-
CONCLUSIONES:
1.- Las células eucarióticas son de mayor
tamaño que las procariótidas y su estructura es
más compleja, y es aquella que posee un núcleo
celular.
2.- Los virus son seres vivientes de tamaño mucho
menor que las células eucarióticas y
procariótidas, y no se les considera como unidades
celulares.
3.- La mayor parte de las células son
microscópicas, pero su tamaño varía en un
rango muy amplio, algunas células bacterianas pueden
apreciarse en un buen microscopio
óptico, y ciertas células animales tienen un
tamaño que permite apreciarlas a simple vista.
4.- El hecho de que una membrana permita el paso de las
moléculas de cierta sustancia depende de la estructura de
aquella y del tamaño y carga eléctrica de las
moléculas.
5.- En los cloroplastos es donde se lleva a cabo la
fotosíntesis, éstos cloroplastos al igual que las
mitocondrias, poseen una membrana interna y una
externa.
6.- En la fotosíntesis las células
transforman la energía luminosa en energía
química, con el aprovechamiento del CO2 de la
atmósfera y la formación de carbohidratos, y
desprendimiento de oxígeno
7.- En la fotorrespiración celular, éstas
utilizan sustratos como la glucosa y producen dióxido de
carbono, esto se lleva a cabo principalmente en sus
mitocondrias.
8- La fermentación va invariablemente
acompañada del desarrollo de microorganismos, y cada tipo
químico de fermentación en particular definido en
función de sus principales productos orgánicos
finales ( por ejemplo la fermentación láctica,
alcohólica o butírica), va acompañada del
desarrollo de un tipo específico de
microorganismo.
9- La fermentación es un proceso menos eficaz que
la respiración aeróbica en cuanto a suministro
energético ya que parte de la energía presente en
la sustancia descompuesta está todavía presente en
los productos orgánicos finales ( por ejemplo, el alcohol o el
ácido láctico) formados de manera
característica en los procesos fermentativos.
10.- Las bacterias se subdividen en un total de 19
partes, cada una de las cuales se distinguen por unos pocos
criterios fáciles de determinas.
11.- La definición más sencilla de la
estructura de un hongo superior es una masa de citoplasma
multinucleda, móvil dentro de un sistema de tubos muy
ramificados que lo encierran.
12.- Los hongos están siempre encerrados dentro
de una pared rígida, no puede atrapar microorganismos
más pequeños. La mayoría de los hongos viven
libres en el suelo o en el agua y obtienen su energía por
respiración o fermentación de materiales
orgánicos solubles presentes en estos
ambientes.
13.- La mejor manera de considerar a los protozoos es
como un conjunto que abarca un cierto número de grupos de
protistas unicelulares eucarióticos ,típicamente
móviles y no fotosintéticas, que probablemente han
derivado, en varios momentos distintos del pasado evolutivo, de
uno u otro grupo de las algas unicelulares
14.- La clasificación primaria de las algas
está basada en propiedades celulares, no del organismo: la
naturaleza química de la pared, si está presente,
los materiales orgánicos de reserva producidos por la
célula; la naturaleza de los pigmentos
fotosintéticos y la naturaleza y disposición de los
flagelos que llevan las células móviles.
15.- La mayoría de las algas son organismos
acuáticos, que habitan las aguas dulces y los
océanos. Estas formas acuáticas son principalmente
de vida libre, pero ciertas algas marinas unicelulares han
establecido relaciones simbióticas duraderas con animales
invertebrados marinos ( por ejemplo, esponjas, corales ) y se
desarrollan dentro de las células del animal
hospedados.
16.-La reproducción de los virus animales en
forma resumida se origina con la adsorción y
penetración del virus animal a una célula
hospedadora con la formación de enlaces covalentes entre
la superficie del virión y los receptores
específicos de la superficie celular, posteriormente el
primer paso en el desarrollo vírico intracelular es la
transcripción y traducción de los genes
víricos.
REFERENCIAS
BIBLIOGRAFICAS
- Ville Claude, Biología, 2° edición,
ed. Mc Graw-Hill, México D.F, 1992, p.78
- Monitor, Enciclopedia Salvat para todos, ed. Salvat,
Barcelona, España,
1965, tomo 4, p. 1317
- IBID (1), p. 82
- IBID (1), p. 83
- IBID (1), p. 79
- IBID (1), p. 80
- Atlas de Biología, Los mecanismos de la vida,
ed. Cultural S.A, Madrid, España, 1992, p.
40
- IBID (7), pp. 46,47
- IBID (1), pp. 5-11
- IBID (1), p. 7
- IBID (1), pp. 113-125
- De Robertis, Biología celular y molecular,
10° edición, ed. Florida, Argentina,
1984, pp. 301-305
- IBID (1), pp. 195,196
BIBLIOGRAFIA:
- Ville Claude, Biología, 2° edición,
ed. Mc Graw-Hill, México D.F, 1992
- Monitor, Enciclopedia Salvat para todos, ed. Salvat,
Barcelona, España, 1965
- Atlas de Biología, Los mecanismos de la vida,
ed. Cultural S.A, Madrid, España, 1992
- De Robertis, Biología celular y molecular,
10° edición, ed. Florida, Argentina,
1984













